по Материалам Международной конференции «Экологическая физиология водных фототрофов: распространение, запасы, химический состав и использование»
V Сабининские чтения 29 ноября 2016 - 29 января 2017
Геохимические условия ландшафта как фактор формирования видового состава макроскопических водорослей в малых реках Байкальского и Амурского бассейнов
Geochemical conditions of the landscape as a factor of formation of the species of macroscopic algae in small rivers of the Baikal and Amur basins
Куклин А.П.
Alex P. Kuklin
Институт природных ресурсов, экологии и криологии СО РАН (г. Чита)
УДК 574.21
Разница в геохимической среде ландшафтов определяет различия в стоке основных биогенных элементов (азота и фосфора). В свою очередь это определяет различия в составе и распределении видов макроскопических водорослей в малых реках Байкальского и Амурского бассейнов.
Ключевые слова: макроводоросли; ландшафт; малая река; биогенные элементы
Введение
Макроскопические водоросли являются обычным компонентом как пресноводных, так и морских экосистем. Они осваивают участки водных объектов, наиболее подверженные гидродинамическому воздействию (прибойная зона озер и морей, речные перекаты). В малых реках с их крайне неустойчивым гидрологическим режимом макроскопические водоросли имеют конкурентное преимущество перед другими первичными продуцентами органического вещества (высшей водной растительностью и микроводорослями фитопланктона и фитобентоса). Преимущество достигается за счет большой скорости роста и особенностей построения тел организмов (нити, слизистые колонии и др.), определяющих их способность осваивать, наряду с благоприятными, и экстремальные участки дна.
Работ, ориентированных на изучение особенностей экологии пресноводных макроскопических форм, немного, что относится не только к территории Верхнеамурского бассейна, но и ко всей России (Чемерис, Бобров, 2008). Незначительная насыщенность видами сообществ, преобладание в их составе видов-космополитов, трудности в систематической идентификации (стерильные формы, определяемые до рода), упор на исследование планктона озер и водохранилищ, а не фитобентоса рек, в целом не способствуют росту числа исследований данной группы организмов. Наибольшее разнообразие макроводорослей отмечается в малых и средних реках – наиболее уязвимом при хозяйственном освоении территории элементе гидросферы.
К настоящему времени макроскопические водоросли в реках Байкальского и Амурского бассейнов на территории Забайкальского края изучены достаточно полно как в видовом отношении (Егорова и др., 2009; Куклин, 2002, 2004, 2006, 2009 а, б, 2011, 2012, 2013 а, б, 2014; Куклин и др., 2002; Оглы, 2011; Оглы, Качаева, 1999), так и во взаимосвязях с другими организмами (Матафонов, Куклин, Матафонов, 2005; Салтанова, Куклин, Макарченко, 2010; Ташлыкова, Куклин, 2014) и окружающей средой (Куклин, Замана, 2011; Kuklin, Zamana, 2009). Это позволяет на основе данных о факторах, обуславливающих пространственное распределение сообществ водорослей в естественных и антропогенно-нарушенных водотоках, прогнозировать видовой состав и особенности развития макроскопических водорослей.
Материалы и методы
В настоящей работе к макроводорослям отнесены низшие водные автотрофные организмы более 2 мм, а также мейоводоросли (0,5–2 мм), образующие слизистые дерновинки и колонии макроскопических размеров.
К категории «самая малая река» и «малая река» в гидрологических справочниках относятся водотоки протяженностью не более 50 км (например: Ресурсы поверхностных вод, 1966). Согласно ГОСТ 17.1.1.02-77 к «малой» относят реку, бассейн которой расположен в одной географической зоне и гидрологический режим ее под воздействием местных факторов может отличаться от свойственного для рек этой зоны. Другой, альтернативной точкой зрения на выделение категории «малая река» является площадь водосбора (Крыленко, Крыленко, Дзагания, 2005). Согласно ГОСТ 19179-73, к малым равнинным рекам относят реки с площадью водосбора меньше или равной 2000 км2 (в засушливых районах – менее 3000 км2). Также к малым равнинным рекам предлагают относить: в зоне избыточного и достаточного увлажнения – реки с площадью водосбора менее 2000 км2, а в лесостепной и степной зонах – реки с площадью водосбора менее 5000 км2 (Методические .., 1986).
Материалы были собраны с 1998 по 2016 гг. на реках Забайкальского края (рис. 1). Сбор и обработка водорослевых образцов проводились согласно общепринятой методике сбора и изучения пресноводных водорослей (Водоросли, 1989). Часть водорослевых образцов изучались в живом виде, прочие – в фиксированном 4%-ным раствором формальдегида. Водоросли изучались с помощью световых микроскопов Биолам Р (ЛОМО, С.- Петербург ) и Nicon Eclipse E200 (Nikon, Япония) под разными увеличениями (от 120 до 400х) в зависимости от размера водорослей исследуемого таксона.
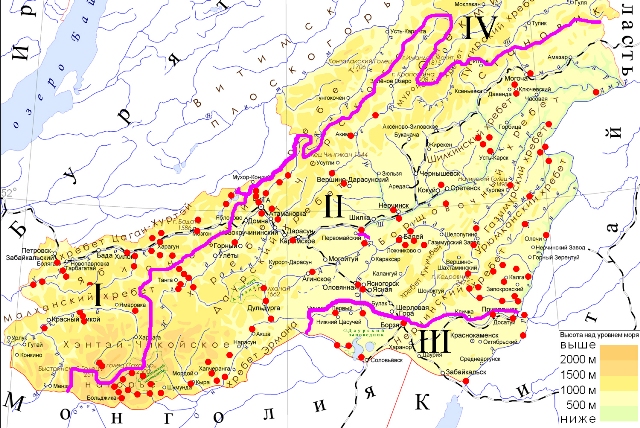
Увеличить карту
Рис. 1. Места сбора проб макроводорослей.
Цифрами обозначены бассейны: I – Байкальский; II - Амурский; III – внутренняя бессточная область; IV – Ленский.
Fig. 1. Locations of sampling of macroalgae.
These numbers refer to basins: I – Baikal; II – Amur; III – enclosed region; IV – Lena.
Результаты и обсуждение
Донные отложения, представленные различными по крупности фракциями, являются неотъемлемым компонентом реки. Горные породы, слагающие донные отложения, согласно (Атлас... , 1997), имеют преимущественно кристаллический состав, что определяет, во-первых, нейтральность горных пород и продуктов их выветривания (пески, дресва) по отношению к растительным организмам. Это позволяет нам не рассматривать химический состав горных пород как фактор, оказывающий влияние на состав и распределение водорослей в пространстве. Во-вторых, кристаллические горные породы обуславливают низкую минерализацию вод малых водотоков Байкальского и Амурского бассейнов. Однако, в сравнении с более водообеспеченными территориями, например реками Уральского хребта (Шубина, 2007), северо-запада России (Чемерис, Бобров, 2008) и Дальнего Востока (Шестеркина и др., 2010), минерализация малых водотоков Байкальского и Амурского бассейнов оказывается немного выше. Воды малых рек, протекающие в ненарушенных антропогенной деятельностью ландшафтах, имеют преимущественно гидрокарбонатный состав. Преобладающими катионами являются кальций и магний. Наименее минерализованы воды рек Хентей-Чикойского нагорья (20–40 мг/л), наиболее минерализованы водотоки, пересекающие Внутреннюю бессточную область, например р. Урулюнгуй (до 500 мг/л). От истоков к устью отмечается рост минерализации. Это связано с испарительным концентрированием и насыщением элементами при растворении горных пород. Известно, что ненасыщенные маломинерализованные воды обладают большей растворяющей способностью (Перельман, 1982).
Исследуемая нами территория на большей своей части дренируется малыми водотоками, что в большой степени обусловлено именно невысоким модулем стока с территории. Максимальных значений модуль стока (10–12 л•с/км2) достигает в высокогорных частях Хентей-Чикойского нагорья. На большей же территории он не превышает 1–4 л•с/км2 (Атлас..., 1997). Таким образом, практически для всех водотоков исследуемой территории можно выделить участки формирования стока и области транзитного перемещения водных масс, а для ряда водотоков, пересекающих бессточную область (или имеющие сток только во влажный период) – территории трансформации (уменьшения) стока. Потери в стоке приводят к перестройкам в составе экосистем, и как следствие отражаются на составе их флоры.
Малые реки Забайкальского края от истока к устью дренируют разные геохимические ландшафты (Ландшафты..., 1977). Геохимический процесс, результатом которого являются потоки веществ, протекает в зависимости от геологических, климатических и орографических условий. Следовательно, особенности ландшафта будут определять состав, направление и скорость потоков веществ. Собственно потоки веществ (например, для водорослей – азота и фосфора), а главное доступных для усвоения их форм, будут принимать участие в формировании состава растительности, т.е. определять разнообразие на видовом и ценотическом уровне. Немаловажным фактором геохимических процессов в ландшафтах рассматриваемых бассейнов выступает мерзлота (рис. 2). Глубокое промерзание почв и медленное оттаивание обуславливают сокращенный период почвообразования, снижение интенсивности микробиологической активности. Это выражается в невысокой гумусоносности и, в силу этого, слабой биогенной аккумуляции элементов в черноземе и их выносом в воды. Известно, что соединения азота и фосфора обладают разной миграционной способностью в водах при разных уровнях рН среды. Так, соединения фосфора более подвижны в кислых условиях среды, а соединения азота в щелочных.
Ландшафты высокогорных тундр имеют ограниченное распространение, на рассматриваемой территории преимущественно в горной части Хентей-Чикойского нагорья. Наибольший модуль стока (10–12 л•с/км2) в совокупности с отсутствием почвенного покрова обуславливают низкую минерализацию (в пределах 20 мг/л) и низкие содержания биогенных элементов.
Таежные ландшафты являются основными стокоформирующими территориями для рек Байкальского и Амурского бассейнов. Модуль стока с территории колеблется в пределах от 6 до 4 л•с/км2. Мерзлота здесь имеет островной, массивно-островной и прерывистый характер распространения. Повышенный промывной режим почв обуславливает невысокую минерализацию вод (до 200 мг/л, чаще в пределах 100 мг/л). В связи с этим богатство вод биогенными соединениями возможно только в двух случаях: когда воды контактируют с верхним слоем почвы и когда в реку поступают воды с заболоченного днища долины. В первом случае это происходит в местах разгрузки подземных вод или тающих вод мерзлоты. Во втором случае это происходит в реках, пересекающих места расширения долин. От истока к устью водный поток принимает не только воды боковых притоков, но и воды подруслового стока, дренирующего в том числе и пойму. Плоские днища заболачиваются, что приводит к развитию восстановительных реакций в условиях переувлажнения и затрудненного доступа кислорода, повышению содержания фосфора в надмерзлотном слое почвы. В кислых водах болотных луж, окружающих водоток, фосфор более подвижен, чем азот. Поступление таких кислых вод в водоток с нейтральными и щелочными водами приводит к осаждению фосфора на дно и вовлечению его в круговорот (рис. 2).
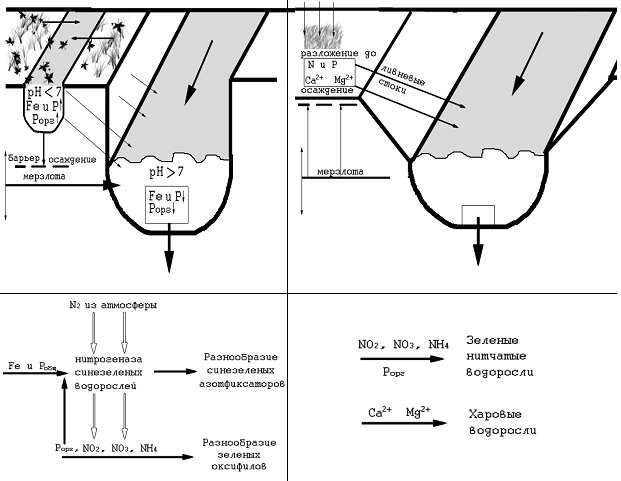
Рис. 2. Геохимические предпосылки разнообразия водорослей.
Fig. 2. Geochemical conditions algae diversity.
Степные ландшафты не являются стокоформирующими в связи с тем, что здесь очень мал модуль стока с территории (не превышает 2 л•с/км2). Мерзлотные процессы развиты слабо, в основном породы талые, возможны перелетки или, на отдельных участках, отмечается спорадическое и редкоостровное распространение многолетнемерзлых пород. Тем не менее, как сообщает Ногина (1964), аккумуляция холода оказывает большое влияние на развитие почв и потоки элементов (в том числе и биогенных) в степных экосистемах. Следует отметить, что большинство малых водотоков в степных ландшафтах эфемерны, т.к. существуют только в периоды обильных атмосферных осадков, или транзитны (например, р. Ага, р. Борзя, р. Урулюнгуй). Эфемерность водотоков не позволяет развиваться организмам с длительным сроком вегетации. Формирование геохимического процесса протекает в условиях отсутствия атмосферного увлажнения в виде осадков большую часть вегетационного сезона. Вода в почве присутствует в виде конденсата влаги в надмерзлотном слое, если таковой имеется (например, солончак) или в слое, аккумулировавшем холод зимой (глубина промерзания до 2 м). Такая вода не может участвовать в стоке, так как она связанная, но она взаимодействует с горными породами, растворяя их. Поднимаясь на поверхность, вода испаряется, оставляя после себя налет солей на почве. В сухой период разложения органического вещества практически не происходит, т.к. нет достаточного количества влаги. Часть сухой прошлогодней ветоши сносится в понижения, которыми являются тальвеги и сухие русла, а также котловины озер. Картина резко меняется в период увлажнения, который в Забайкалье совпадает с наиболее высокими температурами воздуха. В этот период резко интенсифицируются процессы микробного разложения растительной ветоши, как на склонах, так и в руслах рек. Почвы в этот период обогащаются азотом и фосфором (Ногина, 1964). Но также в этот период верхний слой почвы промывается осадками, преимущественного ливневого характера, поэтому биогены интенсивно поступают в водотоки. Туда же поступают и растворимые соли.
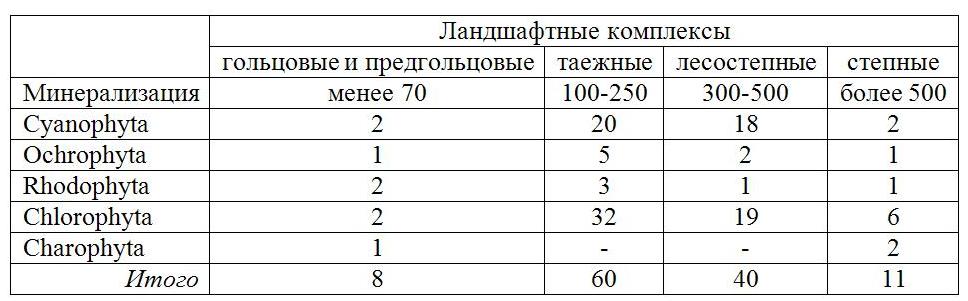
Видовой состав макроводорослей опубликован в наших предыдущих работах (Базарова, Куклин, 2002; Куклин, 2004, 2006, 2009 а,б, 2013 а,б, 2014; Ташлыкова, Куклин, 2014). Выявлено, что количество видов макроводорослей по отделам различается при разной минерализации в реках разных ландшафтных комплексов (табл.1). Различаются и значения фитомассы видов при разной минерализации (табл. 2).
Таблица 1. Минерализация (мг/л) и число видов макроводорослей по отделам
в реках разных ландшафтов
Table 1. Salinity (mg/l) and the number of species of macroalgae in the divisions
in the rivers of different landscapes
Разнообразие макроводорослей в водотоках гольцовых и предгольцовых ландшафтов ограничивается низким количеством питательных веществ, присутствующих в водах. Массовыми видами являются Prasiola fluviatilis (Sommerf.) Aresch (а) и Hydrurus foetidus (Vill.) Trev., реже встречается Tetraspora lubrica (Roth.) Ag.
Наибольшее разнообразие видов отмечено в водотоках, дренирующих таежные и лесостепные ландшафты. Присутствие органического и минерального фосфора стимулирует разнообразие азотфиксаторов. Азот данную группу организмов не лимитирует, так как при наличии фосфора может усваиваться из атмосферы. Например, для широко распространенного на исследуемой территории рода Nostoc характерна скорость азотфиксации в пределах 0,13–0,5 мкгN2•106 клеток (Костяев, 1993). По мнению этого автора, гетероцистные синезеленые водоросли практически все способны усваивать молекулярный азот в аэробных условиях (Костяев, 1993). Нами установлено, что от таежных к степным ландшафтам сокращается доля видов порядка Nostococales (Cyanobacteria), содержащих гетероцисты (Куклин, 2002). В отделе зеленых водорослей происходит сокращение числа видов с пальмелоидной структурой таллома (T. lubrica) или c сильно ослизненными нитями (Chaetophora lobata и Draparnaldia glomerata) и замена их видами с незначительным ослизнением (Ulothrix zonata var. zonata Kütz.) или не ослизняющимися (Cladophora fracta). Организмы с высокой долей воды в талломе – по отношению к органическому веществу – в водотоках таежных зон эффективно конкурируют в борьбе за пространство с другими растительными организмами.
Таблица 2. Минерализация воды (мг/л) и фитомасса (г/м2) макроводорослей
(цит. по Kuklin, Zamana, 2009 с дополнениями)
Table 2. Water salinity (mg/l) and phytomass (g/m2) of macroalgae
(cited by Kuklin, Zamana, 2009 with additions)
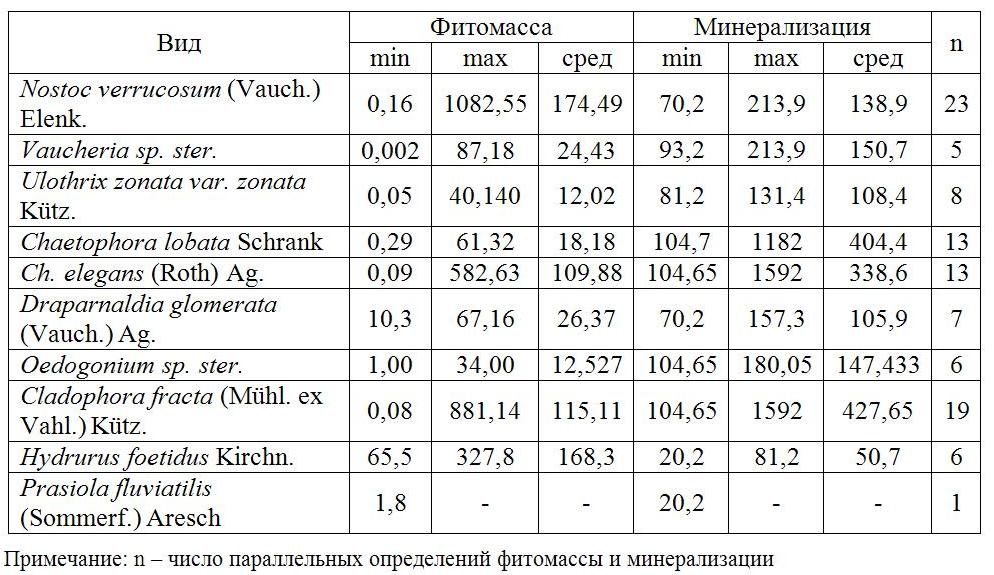
В водотоках степных территорий соотношение азота к фосфору больше, чем в таежных. Это приводит к преобладанию в водотоках лесостепных и степных ландшафтов зеленых нитчатых водорослей. Такой же эффект «цветения нитчатыми» вызывает избыточное поступление соединений азота в реки таежных ландшафтов при антропогенном воздействии, в результате чего доминирование переходит к видам, не характерным для данной территории, например, Hydrodictyon reticulatum (L.) Bory. Этот вид обнаружен нами в р. Ингода у с. Песчанка (в 9,5 км ниже сброса сточных вод г. Чита), в р. Аргунь у с. Молоканка, в которую по данным (Золотые реки, 2012) производится сброс сточных вод г. Чань-Жоу. Этот вид также был отмечен нами и в устье р. Турга (дренажный канал Харанорской ГРЭС) ниже сброса сточных вод с очистных сооружений пос. Ясная и пос. Ясногорск.
Разнообразие макроводорослей в водотоках степных ландшафтов невелико. Оно ограничивается, прежде всего, пересыханием водотоков в летний период. Наличие биогенов создает на короткий период (конец лета – начало осени) благоприятный период для развития нитчатых водорослей (преимущественно Cl. fracta и представителей рода Spirogyra), при минерализации свыше 1 г/л – видов рода Chara. При концентрациях сульфатов в минеральном составе вод свыше 500 мг/л доминантом или субдоминантом сообществ макроводорослей становится Enteromorpha intestinalis (L.) Link. Повышенная минерализация вод препятствует развитию бореальных оксигенных видов.
Таким образом, биогенные элементы через особенности геохимических процессов, протекающих в ландшафтах, оказывают прямое и непосредственное влияние на состав и фитомассу водорослей. Увеличение доли фосфора в воде малых рек таежных ландшафтов ведет к преобладанию гетероцист-содержащих азотфиксирующих синезеленых водорослей (родов Nostoc, Tolypothrix, Rivularia). В лесостепных ландшафтах увеличение доли азота по отношению к фосфору обуславливает обилие оксифильных слизистых форм зеленых водорослей (Ch.lobata, Dr. glomerata, U. zonata var zonata). Повышенная минерализация вод в малых реках степных ландшафтов способствует вегетации E. intestinalis и представителей харовых водорослей.
Благодарности
Работа выполнена по проекту IX.137.1.1 «Биоразнообразие природных и природно-техногенных экосистем Забайкалья (Центральной Азии) как индикатор динамики региональных изменений климата» по программе фундаментальных исследований СО РАН на 2017-2020 гг.
Список литературы
1. Атлас Читинской области и Агинского Бурятского автономного округа. – М: Федеральная служба геодезии и картографии России, 1997. – 48 с.
2. Базарова Б.Б., Куклин А.П. Макрофиты // Ландшафтное и биологическое разнообразие бассейна реки Хилок: опыт изучения и управления. – Новосибирск: Изд-во СО РАН, 2002. – С. 88–99.
3. Водоросли: справочник / Вассер С.П., Кондратьева Н.В., Масюк Н.П. и др. – Киев: Наукова думка, 1989. – С. 130–134.
4. Егорова И.Н., Коновалов М.С., Дударева Н.В. Особенности состава альгофлоры, выявленной в ассоциации с мохообразными Сохондинского биосферного заповедника // Известия ИГУ. Серия: Биология. Экология, 2009. Т.2, №1. С. 8–11.
5. Золотые реки: Выпуск 1. Амурский бассейн. – Владивосток: Всемирный фонд дикой природы (WWF), изд-во Апельсин, 2012 г. – 120 с.
6. Костяев В.Я. Биология, экология и роль азотфиксирующих синезеленых водорослей (цианобактерий) в различных экосистемах. Автореф. дис. докт. биол. наук. – М., 1993. – 20 с.
7. Крыленко И.В., Крыленко В.И., Дзагания Е.В. Что понимать под понятием «малая горная река» и его критерии. – Донецк, 2005. – 14 с. [Электронный ресурс] Проза.ру: сайт. – URL: http://www.proza.ru/2011/07/03/1381.html (дата обращения – 20.11.2016).
8. Куклин А.П. Макрофитные водоросли бассейна реки Аргунь // Аргунские просторы. – 2009 а. – С. 113–119.
9. Куклин А.П. Видовой состав макрофитобентосных сообществ водорослей в притоках реки Хилок // Экосистемы малых рек: биоразнообразие, биология, охрана: тезисы докл. Всерос. конф. (Борок, 16-19 нояб. 2004 г.). – Борок , 2004. – С. 49–50.
10. Куклин А.П. Макроскопические водоросли Sirodotia suecica Kylin., Lemanea fluviatilis Ag., Prasiola fluviatilis (Sommerf.) Aresch, Hydrurus foetidus (Vill.) Trev. в озёрах и реках Забайкалья // Известия Иркутского государственного университета. Серия: Биология. Экология. 2013 б. Т.6, №3. С. 70–76.
11. Куклин А.П. Макроскопические водоросли в бассейне реки Букукун // Записки Забайкальского отделения Русского географического общества. 2013 а. №132. С. 68–75.
12. Куклин А.П. Макроскопические водоросли в водоемах Сохондинского заповедника и охранной зоне // Труды Сохондинского заповедника. Исследования в охранной зоне. Вып. 6. – Чита, 2014. – С. 31–43.
13. Куклин А.П. Макрофитные водоросли – индикаторы загрязнения речной сети приграничных территорий (на примере Кыринского района Забайкальского края) // Природоохранное сотрудничество: Россия, Монголия, Китай. 2011. №2. С. 117–121.
14. Куклин А.П. Макрофитные водоросли в малых реках бассейна реки Хилок // Известия Иркутского государственного университета. Серия: Биология. Экология. 2009 б. Т.2, №1. С. 45–48.
15. Куклин А.П. Состав и особенности развития макроводорослей в притоках реки Онон // Природные ресурсы Забайкалья и проблемы геосферных исследований: материалы науч. конф. (Чита, 12-15 сент.2006 г.). – Чита, ЗабГГПУ, 2006. – С. 83–85.
16. Куклин А.П. Флора макрофитных водорослей (Cyanophyta, Xantophyta, Rhodophyta, Chlorophyta, Charophyta). Аннотированный список флоры и фауны водоемов и водотоков бассейна реки Аргунь. – Новосибирск, 2012. – С. 51–60.
17. Куклин А.П. Экология макрофитных водорослей Восточного Забайкалья (на примере водоемов бассейна р. Хилок). Автореф. дис. канд. биол. наук. – Улан-Удэ, 2002. – 20 с.
18. Куклин А.П., Замана Л.В. Макрофитные водоросли – индикаторы загрязнения речной сети урбанизированных территорий (на примере района г. Читы) // Эволюция биогеохимических систем (факторы, процессы, закономерности) и проблемы природопользования. Геоэкологические, экономические и социальные проблемы природопользования: материалы науч. конф. (Чита, 27-30 сент. 2011 г.). – Чита, 2011. – С. 106–108.
19. Куклин А.П., Матафонов Д.В., Матафонов П.В., Матюгина Е.Б. Биоценозы дна // Ландшафтное и биологическое разнообразие бассейна реки Хилок: опыт изучения и управления. – Новосибирск: Изд. СО РАН, 2002. – С. 140–150.
20. Ландшафты юга Восточной Сибири (карта). М-б 1:1 500 000 / Сост. В.С. Михеев, В.А. Ряшин. – М.: ГУГК, 1977. – 4 л.
21. Матафонов Д.В., Куклин А.П., Матафонов П.В. Консорции в водных экосистемах Забайкалья // Известия Российской академии наук. Сер. биологическая. 2005. №5. С. 592–598.
22. Методические рекомендации по учету влияния хозяйственной деятельности на сток малых рек при гидрологических расчетах для водохозяйственного проектирования / Гос. гидрол. ин-т [Сост. В.Е. Водогрецкий и др.]. – Л.: Гидрометеоиздат, 1986. – 166 с.
23. Ногина Н.А. Почвы Забайкалья. – М., Наука, 1964. – 315 с.
24. Оглы 3.П., Качаева М.И. Биоразнообразие водных экосистем Забайкалья. Каталог водорослей Верхнеамурского бассейна. – Новосибирск: Изд. СО РАН, 1999. – 91 с.
25. Оглы З.П. Фитопланктон разнотипных водных экосистем Восточного Забайкалья. – Чита: ЗабГУ, 2011. – 162 с.
26. Перельман А.И. Геохимия природных вод. – М.: Наука, 1982. – 154 с.
27. Ресурсы поверхностных вод СССР. Основные гидрологические характеристики. Т. 16. Ангаро-Енисейский район. Бассейн оз. Байкал (Забайкалье). – Л.: Гидрометеоиздат, 1966. – 158 с.
28. Салтанова Н.В., Куклин А.П., Макарченко Е.А. К биологии хирономиды Cricotopus (Nostococladius) lygropis Ashe et Murray (DIptera, Chironomidae, orthocladiinae) в реках Забайкальях // Евразиатский энтомологический журнал. 2010. Т.9, №3. С. 393–396.
29. Ташлыкова Н.А., Куклин А.П. Водоросли малых водотоков бассейна р. Онон (Кыринский район, Забайкальский край) // Вестник Северо-Восточного научного центра ДВО РАН. 2014. №2. С. 79–87.
30. Чемерис Е.В., Бобров А.А. Речные криптогамные макрофиты на севере Европейской России // Экосистемы малых рек: биоразнообразие, экология, охрана: лекции и матер. докл. Всеросс. шк.-конф. (Борок, 18-21 нояб. 2008 г.). – Ярославль: изд. OOO «Ярославский печатный двор», 2008. – С. 53–64.
31. Шестеркина Н.М., Таловская В.С., Сиротский С.Е., Шестеркин В.П., Ри Т.Д. Гидроэкологический мониторинг зоны влияния Зейского гидроузла. – Хабаровск: ДВО РАН, 2010. – С. 23–35.
32. Шубина В.Н. Фауна и экология донных беспозвоночных лососевых рек Урала и Тимана. Автореф. дис. докт. биол. наук. – Сыктывкар, 2007. – 33 с.
33. Kuklin A.P., Zamana L.V. Freshwater macroalgae as indicators of water mineralization. 17th International environmental bioindicators conference: Conference theme: Global indicators / International society of environmental bioindicators (ISEBI), Russian society for mass spectrometry, Moscow, Russian Federation, Moscow society of naturalists (MOIP). – M.: Всероссийское масс-спектометрическое о-во, 2009. – P. 68.
Статья поступила в редакцию 29.11.2016
Geochemical conditions of the landscape as a factor of formation of the species of macroscopic algae in small rivers of the Baikal and Amur basins
Alex P. Kuklin
The difference in the geochemical environment of the landscape determine the differences in the flow of biogenic elements (nitrogen and phosphorus). This in turn determines the differences in composition and distribution of macroscopic algae species in small rivers
Keywords: macroalgae; landscape; small river; nutrients
Об авторе
Куклин Алексей Петрович - Kuklin Alex P.
кандидат биологических наук
научный сотрудник, Институт природных ресурсов, экологии и криологии Сибирского отделения РАН, г. Чита, Россия (Institute of Natural Resources, Ecology and Cryology SB RAS, Chita, Russia)
kap0@mail.ru
Корреспондентский адрес: Россия, 672014, г. Чита, ул. Недорезова 16 а, Южный научный центр РАН; телефон (3022) 20-61-97.
ССЫЛКА НА СТАТЬЮ:
Куклин А.П. Геохимические условия ландшафта как фактор формирования видового состава макроскопических водорослей в малых реках Байкальского и Амурского бассейнов // Вопросы современной альгологии. 2017. № 1 (13). http://algology.ru/1099
Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно.
Адрес - info@algology.ru
При перепечатке ссылка на сайт обязательна
К разделу СТАТЬИ
Карта сайта

|
