|
|
|
|
V Сабининские чтения 29 ноября 2016 - 29 января 2017 Унимодальная зависимость скорости роста от объема клеток в культурах черноморских видов микроводорослей
Стельмах Л.В., Мансурова И.М. Lyudmyla V. Stelmakh, Irina M. Mansurova
Институт морских биологических исследований имени А.О. Ковалевского (г. Севастополь)
УДК 582.232:581.121
На основе сопоставления максимальных значений удельной скорости роста и объемов клеток 24 массовых видов черноморских планктонных микроводорослей получена унимодальная зависимость. Выявлены оптимальные размеры клеток, которые позволяют водорослям достигать самых высоких значений удельной скорости роста. При отклонении объема клеток от оптимальных значений, как в сторону его увеличения, так и снижения, удельная скорость роста водорослей постепенно снижается. Ключевые слова: микроводоросли; удельная скорость роста; объем клеток; Черное море.
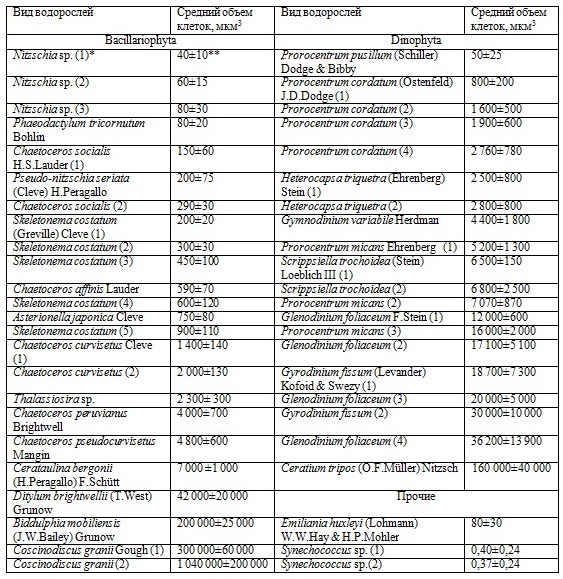
Введение Известно, что прирост биомассы популяции микроводорослей можно характеризовать величинами абсолютной и относительной, или удельной, скорости роста. Прирост биомассы водорослей за сутки (dB/dt) представляет собой абсолютную суточную скорость роста, а ее величина, отнесенная к исходной биомассе водорослей (B), является удельной суточной скоростью роста (m). В течение ряда десятилетий было принято считать, что удельная скорость роста планктонных микроводорослей снижается по мере увеличения объема их клеток (V). Эта связь была описана степенной функцией: m = а´V-b (Banse, 1982, Geider et al., 1986). Однако в последние годы анализ результатов, полученных на морском фитопланктоне, преимущественно олиготрофных районов Мирового океана (Chen, Liu, 2010), и в культурах микроводорослей (Marañón, 2015), позволил показать, что у наиболее мелких клеток, относящихся к пикофитопланктону и нанофитопланктону, скорость роста ниже, чем у более крупных водорослей. В результате связь удельной скорости роста водорослей от объема клеток была представлена в виде унимодальной кривой с максимумом около 100 мкм3. Размерная структура фитопланктона Черного моря отличается от океанического тем, что в нем преобладают более крупные виды, относящиеся преимущественно к нано- и микрофитопланктону. Логично предположить, что связь между объемом клеток и удельной скоростью роста массовых видов черноморских микроводорослей может отличаться от полученных ранее зависимостей. Основную биомассу черноморского фитопланктона создают диатомовые и динофитовые водоросли (Георгиева, 1993), тогда как кокколитофориды и пикопланктон, включая цианобактерии могут доминировать по численности в отдельные периоды года (Kopuz et al., 2012, Stelmakh, Georgieva, 2014). Массовыми видами, характерными для планктона Черного моря, являются представители диатомовых водорослей (15 видов), динофитовых (8 видов), а также кокколитофорида Emiliania huxleyi (Lohmann) W.W.Hay & H.P.Mohler и цианобактерии рода Synechococcus (табл. 1).
Таблица 1. Исследованные виды водорослей и объем их клеток Table 1.The studied species of algae and the volume of their cell
Примечание: 1) * – номер в скобках использован для культур одного и того же вида, выделенных из различных районов Черного моря; 2) ** – приводится стандартное отклонение
Цель настоящей работы заключалась в том, чтобы на основе обобщения данных, полученных на культурах массовых видов черноморских микроводорослей, описать зависимость удельной скорости роста от объема клеток и выявить оптимальные размеры микроводорослей, которые обеспечивают их наибольшую удельную скорость роста в условиях лабораторной культуры.
Материалы и методы В основу работы положены результаты, полученные в основном авторами данной работы на альгологически чистых культурах 24 черноморских видов планктонных микроводорослей, содержащихся в коллекции отдела экологической физиологии водорослей ИМБИ (г. Севастополь). Некоторые виды водорослей представлены несколькими культурами в количестве от двух до пяти, выделенными из природного фитопланктона разных районов прибрежных вод Черного моря в различные годы (поэтому культуры одного и того же вида различаются по среднему объему клеток). Всего в работе было использовано 47 культур микроводорослей. Результаты по максимальной скорости роста для диатомовой водоросли Phaeodactylum tricornutum представлены в работе (Шоман, Акимов, 2013). В анализ были включены данные по объемам клеток и максимальной удельной скорости роста, измеренным в экспоненциальной фазе роста культур при световом насыщении роста, которое для исследованных видов в условиях непрерывного искусственного освещения лампами дневного света находилось в диапазоне от 80 до 200 мкЭ×м-2×сек-1. Температура питательной среды в экспериментах составляла для диатомовых водорослей 15–19°С, для динофитовых и прочих видов – 19–22°С, что соответствовало температурному оптимуму для роста этих видов водорослей. Водоросли культивировали в колбах объемом 250 мл в питательной среде f/2, состав которой представлен в работе (Guillard, Ryther, 1962). Удельную скорость роста культур рассчитывали по формуле, представленной в работе (Заика, 1972) на основе суточного прироста численности клеток или органического углерода, либо по увеличению оптической плотности при длине волны 675 нм, соответствующей максимуму поглощения хлорофилла а в красной области: µ = ln(N2)/ln(N1), где N1, N2 – начальная численность клеток и их количество через сутки (кл/л), или исходная концентрация углерода (мкг/л) и его количество через сутки, либо начальная и конечная величины оптической плотности (условные единицы). Оптическую плотность измеряли с помощью фотоэлектроколориметра КФК-3 в кювете объмом 5 мл в трех повторностях. Численность клеток и их линейные размеры определяли под световым микроскопом ZEISS Primo Star. Общую численность клеток учитывали в капле объемом 0,1 мл в трех повторностях. Для определения линейных размеров клеток использовали от 30 до 50 клеток в зависимости от степени размерной гетерогенности культуры. Объем клеток водорослей (Vкл) рассчитывали на основе принципа геометрического подобия по формулам, представленным в работе (Брянцева, 2005). Эту величину для каждой культуры рассчитывали как среднее арифметическое значение, определенное для всей выборки, а также стандартное отклонение от средней. Содержание органического углерода и азота в клетках водорослей было определено с помощью CHN-анализатора (Grasshoff et al., 1983). Измерения проводили в трех повторностях. На основе полученных значений рассчитывали величину атомарного отношения между органическим азотом и углеродом. Интенсивность света измеряли люксметром Ю-116. Для перехода от освещенности в люксах к энергетическим единицам использовался коэффициент перехода 1000 лк = 17,2 мкЭ×м-2×с-1 (Парсонс, 1982). Построение графиков осуществляли с помощью программы Golden Software Grafer 2001, а расчеты исследуемых параметров и статистическую обработку проводили в программе Microsoft Excel 2007. Расчеты коэффициентов степенных уравнений, отражающих зависимость удельной скорости роста водорослей от объема их клеток, а также коэффициент детерминации (R2) и уровень значимости (р), рассчитывали с помощью программы Sigma Plot 2001 для Windows .
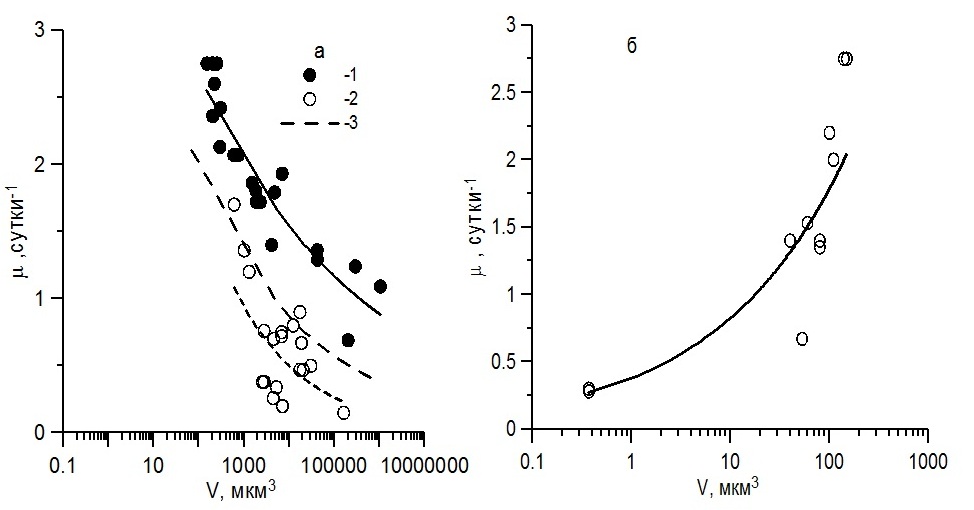
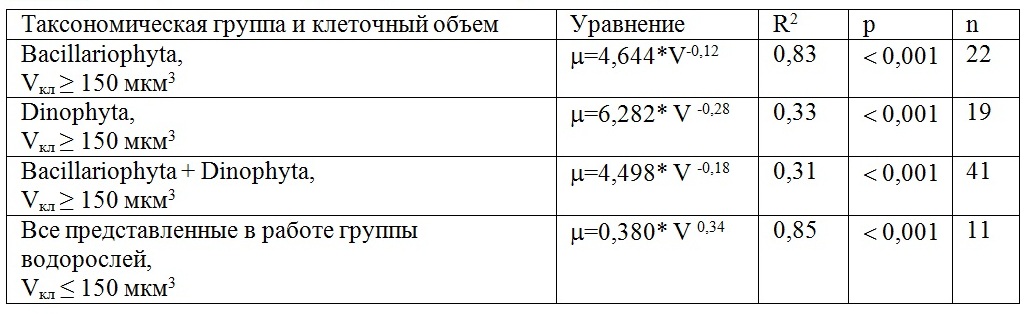
Результаты и обсуждение Для всех живых организмов от бактерий до крупных растений и животных связь удельной скорости метаболизма с массой тела или объемом обычно представляют в виде степенной функции, для которой общепринятым считается степенной коэффициент, равный -0,25 (Chen, Liu, 2010). Исследования на культурах микроводорослей с объемом клеток около 100 мкм3 и более показали, что в пределах одной таксономической группы по мере увеличения размеров клеток максимальная удельная скорость роста водорослей снижается. Эти зависимости получены преимущественно для диатомовых водорослей (Banse, 1982; Finkel, 2001; Стельмах и др., 2010) и гораздо реже для динофитовых (Banse, 1982, Стельмах и др., 2010). По мнению исследователей, при оптимальных световых и температурных условиях, а также достаточном количестве биогенных веществ в среде снижение скорости роста микроводорослей по мере увеличения объема клеток обусловлено, прежде всего, ослаблением внутриклеточной транспортировки биогенных веществ и снижением удельного содержания хлорофилла а в клетках (Marañón, 2015). В Черном море в фитопланктоне представлены виды водорослей, имеющие объем клеток от 0,37–0,40 мкм3 до 106 мкм3 (табл. 1). Среди диатомовых наиболее мелкие размеры имеют виды рода Nitzschia (Vкл = 40–80 мкм3) и вид Phaeodactylum tricornutum, а среди динофитовых – Prorocentrum рusillum (Vкл = 50 мкм3); к мелкоклеточным видам водорослей также можно отнести кокколитофориду Emiliania huxleyi (80 мкм3). Наименьший размер клеток среди рассмотренных нами черноморских видов водорослей отмечен у цианобактерии Synechococcus sp. (объем клеток менее 1 мкм3). Наиболее крупные клетки отмечены у диатомовой водоросли Сoscinodiscus granii, а из динофитовых – у Сeratium tripos (с объемом клеток 1 040 000 мкм3 и 160 000 мкм3 соответственно). Выполненные ранее исследования на культурах показали, что при оптимальных условиях наиболее высокие значения скорости роста характерны для диатомовых водорослей, достигающие 1,40–2,70 сутки-1 (Стельмах и др., 2010). Тогда как у семи культур динофитовых водорослей, таких как P. pussilum, P. cordatum, P. micans, S. trochoidea, G. fissum, G. foliaceum и H. triquetra, представленных в данной работе, значения максимальной скорости роста были существенно ниже, изменяясь от 0,34 до 1,36 сутки-1 (Мансурова, 2013). Это обусловлено тем, что удельное содержание хлорофилла а в клетках динофитовых водорослей, как правило, в 2–3 раза меньше, чем у диатомовых видов (Стельмах, 1982; Finenko et al., 2003). Относительные значения дыхания у динофитовых водорослей выше, чем у диатомовых, что подтверждается экспериментами на культурах и полевыми данными. Например, в обзорной статье Гейдера и Осборна (Geider, Osborn, 1989) показано, что в активно растущих культурах морских микроводорослей при температуре 18–22оС интенсивность «темнового» дыхания изменяется в широком диапазоне: от 1 до 59% от валового значения скорости фотосинтеза. Максимальное среднее значение этого параметра (35%) получено для динофитовых водорослей, минимальное (7%) – для цианобактерий. Промежуточная величина (в среднем 14%) наблюдалась у диатомовых водорослей. Различия в значениях максимальной скорости роста микроводорослей (µмакс) приводят к необходимости построения зависимостей этого параметра от объема клеток (Vкл) отдельно для диатомовых и динофитовых водорослей. Как видно, связь µмакс с Vкл для диатомовых водорослей с объемом клеток от 150 мкм3 и выше хорошо описывается обратной степенной зависимостью (рис. 1а) с высоким значением коэффициента детерминации, который равен 0,83 (табл. 2). Подобная связь наблюдалась и для динофитовых водорослей. Однако степенной коэффициент этой зависимости по абсолютной величине был приблизительно в 2 раза выше, чем у диатомовых водорослей. Иными словами, в отличие от диатомовых водорослей скорость роста динофитовых гораздо быстрее убывает по мере увеличения объема клеток, хотя коэффициент детерминации этой связи был существенно ниже и составил 0,33 (табл. 2). По нашим результатам видно, что построение общих зависимостей скорости роста от объема клеток для водорослей, принадлежащих к двум различным таксономическим группам, приводит к снижению коэффициента детерминации, а значит к ослаблению связи между указанными параметрами, что ранее было показано и другими авторами (например, Tang, 1996). Ослабление связи между µмакс и Vкл происходит также при отклонении условий среды от оптимальных.
Рис. 1. Зависимость между максимальной удельной скоростью роста и объемом клеток водорослей для видов с объемом клеток более 150 мкм3 (а): для диатомовых (1), динофитовых (2) и общая для обеих групп водорослей (3); связь между скоростью роста и объемом клеток для видов водорослей с объемом клеток 150 мкм3 и менее для всех представленных в работе таксономических групп (б) Fig.1. The relationship between the specific growth rate and volume for algal cells with volume more 150 mkm3 (a): for the diatoms (1), dinoflagellates (2) and common to both groups of algae (3); relationship between the growth rate and volume for algal cells with volume 150 mkm3 or less for all taxonomic groups of algae (b)
Таблица 2. Уравнения, отражающие зависимость удельной скорости роста водорослей Table 2. The equation showing the dependence of specific algal growth rate from the volume of their cells
Примечание: R2 – коэффициент детерминации, p – уровень значимости, n – объем выборки
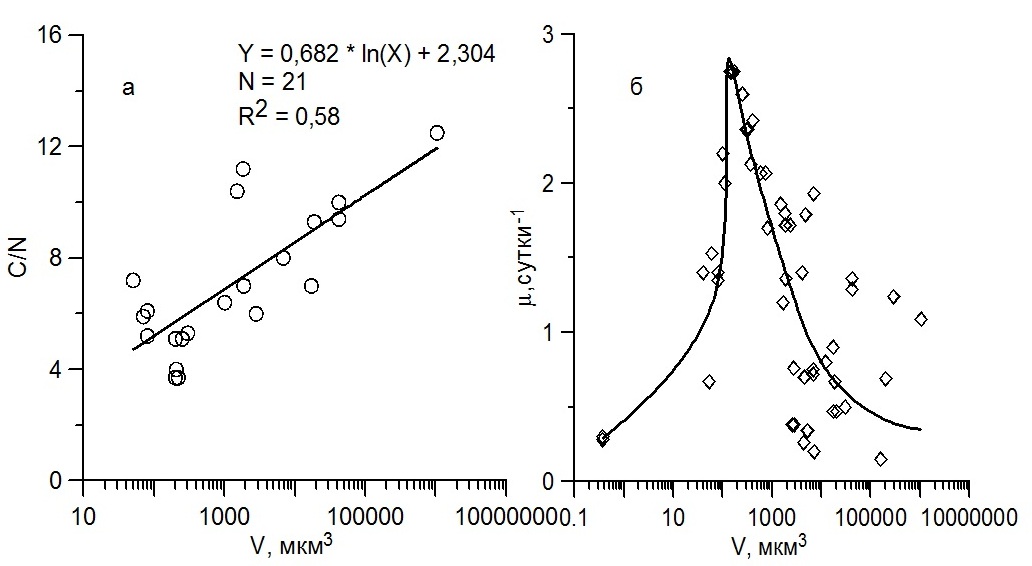
Так, при низких значениях интенсивности света или при недостатке биогенных веществ в среде, связь между удельной скоростью роста и объемом клеток водорослей даже в пределах одной таксономической группы выражена значительно слабее, чем в условиях светового насыщения роста и достаточном количестве питательных веществ (Finkel, 2001). При оптимальных условиях среды удельная скорость роста черноморских видов микроводорослей, представленных разными таксономическими группами, у которых объем клеток был ≤ 150 мкм3, связана степенной зависимостью с высоким значением коэффициента детерминации (R2 = 0,85), однако степенной коэффициент имеет положительный знак и равен 0,34 (рис. 1б). Следовательно, для наиболее мелких водорослей по мере увеличения объема клеток удельная скорость роста возрастает. Такая зависимость обусловлена, вероятно, тем, что у самых мелких водорослей содержание азота по отношению к органическому углероду существенно выше, чем у крупных водорослей (рис. 2а). Как считают некоторые авторы (например, см. Raven, 1998), в мелких клетках за счет более высокого содержания азота и его соединений, таких как нуклеиновые кислоты и мембранные белки, свободного пространства для ферментов и других структур, участвующих в биосинтезе, меньше, чем в относительно крупных клетках. По мере увеличения объема клеток от 0,1 до 100 мкм3 свободное пространство для структур, участвующих в биосинтезе, внутри клеток возрастает, что приводит к увеличению их скорости роста (Marañón, 2015).
Рис. 2. Связь между атомарным отношением органического углерода к азоту (С/N) в клетках и их объемом у диатомовых и динофитовых видов водорослей (а), зависимость между максимальной удельной скоростью роста и объемом клеток для всех размерных групп водорослей разной таксономической принадлежности (б) Fig.2. The relationship between the atomic ratio of organic carbon to nitrogen (C/N) in the cells to their volume for dinoflagellates and diatom algae species (a), the relationship between the specific growth rate and cell volume for all size groups and algae taxa (b)
Обобщение всех данных, полученных нами, позволило представить связь между максимальной удельной скоростью роста массовых видов черноморских микроводорослей и их клеточным объемом в виде единой унимодальной зависимости (рис. 2б). Как видно, наибольшие значения удельной скорости роста (2,5–2,7 сутки-1) наблюдались для клеток, объем которых составлял 150–200 мкм3, т.е. у водорослей с объемом клеток в 1,5–2 раза больше, чем по данным, полученным ранее для схожих условий (по освещенности, температуре и биогенным веществам) (Marañón et al., 2013). Такими являются виды диатомовых водорослей C. socialis и S. сostatum, которые вызывают периодические «цветения воды» в прибрежных водах Черного моря. Первый вид отсутствовал в выборке, использованной в работе наших зарубежных коллег, а второй вид имел объем клеток более 200 мкм3. Это, вероятно, повлияло на положение экстремального значения на графике связи между удельной скоростью роста и объемом клеток водорослей, представленной этими авторами (Marañón et al., 2013). Как следует из полученной нами зависимости, средние значения удельной скорости роста (около 1,5 сутки-1) наблюдались как у мелких водорослей (Vкл ≈ 60–80 мкм3), так и у крупных (1 000-20 000 мкм3). Эти величины скорости роста также достаточны для интенсивного развития водорослей. Однако «цветение воды» среди представленных мелких видов дает только кокколитофорида E. huxleyi, которая, как известно, слабо потребляется микрозоопланктоном (Olson, Strom, 2002). Среди крупных видов интенсивное развитие в Черном море отмечено для диатомовых C. curvisetus и С. bergonii, которые также слабо потребляются микрозоопланктоном (Stelmakh, Georgieva, 2014). Самые низкие величины удельной скорости роста (около 0,20–0,30 сутки-1) выявлены для водорослей с объемом клеток около 0,40 и 200 000 мкм3. С одной стороны, это цианобактерия Synechococcus sp., с другой – динофитовая водоросль C. tripos. Исключение составил самый крупный вид – диатомовая водоросль С. granii, у которой объем клеток был более 106 мкм3, при этом, максимальная удельная скорость роста этого вида была достаточна высокой и составила 1,2 сутки-1.
Заключение Связь максимальных значений удельной скорости роста черноморских микроводорослей с объемом их клеток, который варьировал в пределах семи порядков величин, представлена в виде унимодальной зависимости. Ее максимум (2,5–2,7 сутки-1) отмечен для клеток диатомовых водорослей C. socialis и S. сostatum с объемом клеток 150–200 мкм3. При отклонении клеточного объема от оптимальных значений, как в сторону его увеличения, так и снижения, удельная скорость роста микроводорослей постепенно уменьшается. Самые низкие величины (около 0,20–0,30 сутки-1) выявлены у наиболее мелких клеток, имеющих объем около 0,40 мкм3 и принадлежащих цианобактерии Synechococcus sp., а также у крупных клеток динофитовой водоросли C. tripos с объемом около 200 000 мкм3.
Благодарности. Авторы статьи выражают искреннюю благодарность Кожемяка А.Б. за помощь в выполнении работы. Работа выполнена в рамках государственного задания по направлению «Изучение механизмов адаптации, трансформации и эволюции морских и океанических экосистем в условиях климатических изменений и антропогенного влияния» (№ 1001-2014-0013).
Список литературы 1. Брянцева Ю.В., Лях А.М., Сергеева А.В. Расчет объемов и площадей поверхности одноклеточных водорослей Черного моря. – Севастополь: Препр. ИнБЮМ НАН Украины, 2005. – 25 с. 2. Георгиева Л.В. Видовой состав и динамика фитоцена // Планктон Черного моря. – Киев: Наук. думка, 1993. – С. 31–55. 3. Заика В. Е. Удельная продукция водных беспозвоночных – Киев: Наук. думка, 1972. – 143 с. 4. Мансурова И.М. Влияние света на удельную скорость роста динофитовых водорослей Чёрного моря // Морск. экол. журн. 2013. Т.12, № 4. С. 73–78. 5. Парсонс Т.Р., Такахаши М., Харгрей В. Биологическая океанография. – М.: Легкая и пищ. пром-сть, 1982. – С. 89–91. 6. Стельмах Л.В., Куфтаркова Е.А., Акимов А.И., Бабич И.И., Кожемяка А.Б. Использование переменной флуоресценции хлорофилла in vivo для оценки функционального состояния фитопланктона // Системы контроля окружающей среды. 2010. №13. С. 263–268. 7. Стельмах Л.В. Суточные изменения относительного содержания хлорофилла «a» в культурах морских планктонных водорослей // Экология моря. 1982. №1. С. 68–72. 8. Шоман Н.Ю., Акимов А.И. Влияние фотоадаптации на удельную скорость роста и соотношение органического углерода к хлорофиллу а у диатомовой водоросли Phaeodactylum tricornutum // Морской экологический журнал. 2013. Т.12, №4. С. 97–103. 9. Banse K. Cell volumes, maximal growth rates of unicellular algae and ciliates, and the role of ciliates in the marine pelagial // Limnol. Oceanogr. 1982. V.27. P. 1059–1071. 10. Сhen B., Liu H. Relationships between phytoplankton growth and cell size in surface oceans: Interactive effects of temperature, nutrients and grazing // Limnol. Oceanogr. 2010. V.55. P. 965–972. 11. Finenko Z.Z., Hoepffner N., Williams R., Piontkovski S.A. Phytoplankton carbon to chlorophyll a ratio: response to light, temperature and nutrient limitation // Mar. Ecol. J. 2003. V.2, №2. P. 40–64. 12. Finkel Z.V. Light absorption and size scaling of light-limited metabolism in marine diatoms // Limnol. Oceanogr. 2001. V.46. P. 86–94. 13. Geider R.J., Platt N., Raven I.A. Size dependence of growth and photosynthesis in diatoms: a synthesis // Mar. Ecol. Prog. Ser. 1986. Vol.30. P. 93–104. 14. Geider R., Osborn B.A. Respiration and microalgal growth: a review of the quantitative relationship between dark respiration and growth // New. Phytol. 1989. V.112. P. 327–341. 15.Grasshoff K., Ehrhardt M., Kremling K. Methods of seawater analysis. – Weinheim, Germany: Verlag Chemie, 1983. – 419 p. 16. Guillard R.R.L., Ryther J.H. Studies of marine planktonic diatoms: I. Cyclotella nana Hustedt, and Detonula confervacea (Cleve) Gran // Can. J. Microbiol. 1962. V.8, №2. P. 229–239. 17. Kopuz U, Feyzioglu A. M., Agirbas E. Picoplankton Dynamics during Late Spring 2010 in the South-Eastern Black Sea // Turk. J. Fish. Aquat. Sci. 2012. V.12. Р. 397–405. 18. Marañón E., Cermeño P., Lόpez–Sandoval D.C. et al. Unimodal size scaling of phytoplankton growth and the size dependence of nutrient uptake and use // Ecology Letters. 2013. V.16. P. 371–379. 19. Marañón E. Cell size as a key determinant of phytoplankton metabolism and community structure // Annu. Rev. Mar. Sci. 2015. V.7. P. 241–264. 20. Olson M.B., Strom S.L. Phytoplankton growth, microzooplankton herbivory and community structure in the southeast Bering Sea: Insight into the formation and temporal persistence of an Emiliania huxleyi bloom // Deep-Sea Res. 2002. V.49 (II). P. 5969–5990. 21. Raven J.A. The twelfth Tansley Lecture. Small is a beautiful: the picoplankton // Funct. Ecol. 1998. V.12. P. 503–513. 22. Stelmakh L.V., Georgieva E.Yu. Microzooplankton: the trophic role and involvement in the phytoplankton loss and bloom-formation in the Black Sea // Turkish Journal of Fish. Aquat. Sciences. 2014. V.14. P. 955–964. 23. Tang E.P.Y. Why do dinoflagellates have lower growth rates // J. Phycol. 1996. V.32. P. 80–84. Статья поступила в редакцию 18.12.2016
Unimodal dependence of the growth rate from cell volume in the cultures of the Black Sea microalgae Lyudmyla V. Stelmakh, Irina M. Mansurova The unimodal dependence was obtained on the basis of comparison of the maximum specific growth rate values and cell volumes of 24 dominant planktonic species of microalgae from the Black Sea. The optimal cell sizes allowed algae to reach the highest specific growth rate values were found. When cell volume deviates from the optimal values in the direction of its increase or decrease, the specific growth rate will gradually reduce. Keywords: microalgae; specific growth rate; cell volume; the Black Sea.
Об авторах
Стельмах Людмила Васильевна – Stelmakh Lyudmyla V. кандидат биологических наук ведущий научный сотрудник, Институт морских биологических исследований им. А.О. Ковалевского РАН, Севастополь, Росия (The A. O. Kovalevsky Institute of Marine Biological Research of RAS, Sevastopol, Russia), Отдел экологической физиологии водорослей lustelm@mail.ru Мансурова Ирина Мяулитовна – Mansurova Irina M. младший научный сотрудник, Институт морских биологических исследований им. А.О. Ковалевского РАН, Севастополь, Росия (The A. O. Kovalevsky Institute of Marine Biological Research of RAS, Sevastopol, Russia), Отдел экологической физиологии водорослей ira.mansurova2013@yandex.ua Корреспондентский адрес: 299011 г. Севастополь, пр. Нахимова, 2, Институт биологии южных морей им. А.О. Ковалевского; тел.: +7 (8692) 54-41-10.
ССЫЛКА НА СТАТЬЮ: Стельмах Л.В., Мансурова И.М. Унимодальная зависимость скорости роста от объема клеток в культурах черноморских видов микроводорослей // Вопросы современной альгологии. 2017. № 1 (13). URL: http://algology.ru/1101
Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно.
При перепечатке ссылка на сайт обязательна
На ГЛАВНУЮ
К разделу ОБЗОРЫ, СТАТЬИ И КРАТКИЕ СООБЩЕНИЯ. 
|
|||
|
| ||