|
|
|
|
по Материалам Международной конференции «Экологическая физиология водных фототрофов: распространение, запасы, химический состав и использование» VI Сабининские чтения 1 ноября - 15 декабря 2017 г. Функциональное состояние и некоторые структурные характеристики морских планктонных водорослей при разном уровне обеспеченности биогенными веществами
Стельмах Л.В. Lyudmyla V. Stelmakh
Институт морских биологических исследований им. А.О. Ковалевского (Севастополь, Россия)
УДК [582.261.1+582.276]:546.171.6(262.5)
На культурах 10 видов черноморских планктонных водорослей исследовано влияние содержания биогенных веществ в среде на изменчивость внутриклеточного C/N отношения, удельной скорости роста культур, относительной переменной флуоресценции хлорофилла (Fv/Fm) и отношения C/Хл а. При изменении этих параметров наиболее тесная корреляция между ними наблюдается в пределах одного вида водорослей, а для разных видов она ослабевает или отсутствует. Ключевые слова: микроводоросли; удельная скорость роста; относительная переменная флуоресценция; внутриклеточное содержание азота; отношение между органическим углеродом и хлорофиллом а.
Введение Одно из основных направлений современных гидробиологических исследований связано с оценкой современного состояния морских экосистем, путей их трансформации и развития в условиях глобального потепления и усиления антропогенного воздействия. Важной составляющей этих исследований является оценка функционального состояния первичного звена экосистем – фитопланктона. В практике гидробиологических исследований для оценки функционального состояния морского фитопланктона чаще всего используют такие показатели, как удельная скорость роста и относительная переменная флуоресценция хлорофилла in vivo, а также отношение между органическим углеродом и хлорофиллом а в клетках водорослей (Маторин и др., 1996; Стельмах, Бабич, 2003; Стельмах и др., 2010; Geider, 1987). Однако при изменении абиотических условий показатели функционального состояния фитопланктона не всегда коррелируют между собой (Финенко и др., 2017). Мы полагаем, что это обусловлено, прежде всего, периодической сменой видов в фитопланктонном сообществе и разным откликом названных выше параметров в ответ на изменение не только условий среды, но и перестройку видового состава фитопланктона. В качестве методологической основы для подтверждения нашего предположения могут служить исследования на отдельных видах микроводорослей, выращенных при разном уровне их обеспеченности биогенными веществами. Цель настоящей работы – исследовать изменчивость удельной скорости роста, относительной переменной флуоресценции хлорофилла in vivo и отношения между органическим углеродом и хлорофиллом а в культурах массовых представителей черноморских планктонных водорослей при разном уровне содержания биогенных веществ в среде.
Материалы и методы Объектом исследования послужили альгологически чистые культуры диатомовых водорослей Skeletonema costatum (Greville) Cleve, Chaetoceros socialis H.S.Lauder, C. curvisetus Cleve и Ditylum brightwellii (T.West) Grunow, а также динофитовых водорослей Prorocentrum micans Ehrenberg, P. cordatum (Ostenfeld) J.D.Dodge, P. pusillum (Schiller) Dodge & Bibby, Glenodinium foleaceum F.Stein, Gyrodinium fissum (Levander) Kofoid & Swezy и Heterocapsa triquetra (Ehrenberg) Stein, представленных в коллекции Отдела экологической физиологии водорослей ИМБИ им. А.О. Ковалевского РАН. Культуры выращивали в колбах объемом 250 мл в питательной среде f/2 (Guillard, Ryther, 1962) в накопительном режиме в течение 5–7 дней. Световые условия соответствовали оптимальным для роста водорослей. Они создавались за счет непрерывного искусственного освещения лампами дневного света, интенсивность которого находилась в диапазоне от 100 до 200 мкЭ×м-2×сек-1. Температура питательной среды в экспериментах составляла для диатомовых водорослей 15–19°С, для динофитовых – 19–22°С, что соответствовало температурному оптимуму для роста этих видов водорослей. После прохождения логарифмической фазы роста и достижения стационарной фазы водоросли переносили в стерильную морскую воду без добавления питательных веществ и культивировали при таких же световых и температурных условиях в течение 3–5 дней, что приводило к постепенному снижению структурных и функциональных параметров исследуемых культур до минимальных значений. Эти параметры микроводорослей (внутриклеточное атомарное отношение С/N, отношение между органическим углеродом и хлорофиллом а, относительная переменная флуоресценция хлорофилла и удельная скорость роста) измеряли в логарифмической фазе роста культур при оптимальных по биогенным веществам условиях, при их недостатке в условиях стационарной фазы роста, а также выращивания водорослей на обедненной морской воде, в которой минеральные соединения азота, кремния и фосфора практически отсутствовали. Удельную скорость роста культур рассчитывали по формуле, представленной в работе (Заика, 1972) на основе суточного прироста численности клеток или органического углерода, либо по увеличению оптической плотности на длине волны 675 нм, соответствующей максимуму поглощения хлорофилла а в красной области: µ = ln(N2)/ln(N1), где N1, N2 – начальная численность клеток и их количество через сутки (кл/л), либо исходная концентрация углерода (мкг/л) и его количество через сутки, либо начальная и конечная величины оптической плотности (условные единицы). Оптическую плотность измеряли с помощью фотоэлектроколориметра КФК-3 в кювете объемом 5 мл в трех повторностях. Численность клеток и их линейные размеры определяли под световым микроскопом ZEISS Primo Star. Общую численность клеток учитывали в капле объемом 0,1 мл в трех повторностях. Относительная ошибка определения численности клеток не превышала 10%. Для определения содержания органического углерода и азота в клетках отбирали пробы объемом 100–150 мл в двух повторностях, фильтровали через фильтры GF/F и сжигали в СНN-анализаторе (Grasshoff et al., 1983). Относительная ошибка определения азота и углерода составляла 10–12%. Концентрацию хлорофилла а в микроводорослях измеряли в ацетоновых экстрактах флуориметрическим методом (Holm-Hansen et al., 1965). Подробная методика представлена в статье (Стельмах, Бабич, 2003). Измерения относительной переменной флуоресценции хлорофилла (Fv/Fm) проводили на двухвспышечном флуориметре изготовленном в Отделе экологической физиологии водорослей ИМБИ и подробно описанном ранее (Стельмах и др., 2010). Ошибка определений величины Fv/Fm не превышала 5%. Считается, что отношение Fv/Fm соответствует потенциально возможной эффективности использования энергии света реакционными центрами фотосистемы 2 – РЦ ФС2 (Маторин и др., 1996). Так как на свету активность реакционных центров снижается, перед началом измерений флуоресцентных сигналов культуры микроводорослей выдерживали в темноте в течение 30 мин для того, чтобы все реакционные центры ФС2 перешли полностью в открытое состояние. Интенсивность света измеряли люксметром Ю-116. Для перехода от освещенности в люксах к энергетическим единицам использовался коэффициент перехода 1000 лк = 17,2 мкЭ×м-2×с-1 (Парсонс, 1982). Построение графиков осуществляли с помощью программы Golden Software Grafer 3, а расчеты исследуемых параметров и статистическую обработку проводили в программе Microsoft Excel 2007.
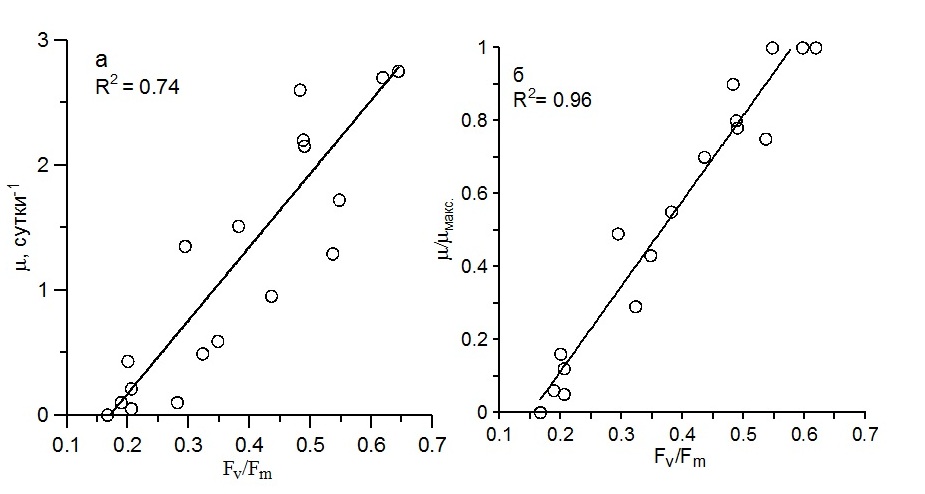
Результаты и обсуждение Прирост биомассы водорослей за сутки (dB/dt) представляет собой абсолютную суточную скорость роста. А ее величина, отнесенная к среднесуточной биомассе водорослей (B), является удельной суточной скоростью роста (Заика, 1972). Определение этого параметра в морском фитопланктоне включает достаточно трудоемкую и длительную процедуру подготовки проб для их суточной экспозиции и последующего анализа (Стельмах и др., 2010). Тогда как относительная переменная флуоресценция хлорофилла in vivo применяется для быстрой диагностики функционального состояния морского фитопланктона и отдельных видов водорослей (Kolber, Falkowski, 1993; Маторин и др., 1996). Переменная флуоресценция связана с активностью реакционных центров фотосистемы 2 (ФС2), где осуществляются световые реакции фотосинтеза. Эффективность этих реакций зависит от факторов среды, и прежде всего, от света и содержания биогенных веществ в воде (Маторин и др., 1996; Berges, Falkowski, 1998). Последние, как известно, оказывают большое влияние на такой функциональный показатель фитопланктона, как удельная скорость роста. Можно предположить, что между скоростью роста и переменной флуоресценцией микроводорослей существует количественная связь. Для выявления этой связи были использованы три вида диатомовых водорослей S. costatum, D. brightwellii и C. socialis. Было получено, что при оптимальных для роста световых и температурных условиях удельная скорость роста в накопительных культурах связана с величиной относительной переменной флуоресценции линейной зависимостью с коэффициентом детерминации, составившим 0,74 (рис. 1а). Максимальные значения обоих параметров, представленные на графике, наблюдались в фазе экспоненциального роста культур при достаточном уровне биогенных веществ. Минимальные величины отмечены при осуществлении их роста на обедненной биогенными веществами морской воде, промежуточные – в стационарной фазе роста культур, когда содержание биогенных веществ ниже оптимального и начинает ограничивать функционирование водорослей. Как видно, при одном и том же значении относительной переменной флуоресценции удельная скорость роста различается в 2–5 раз. Это обусловлено тем, что в экспериментах использованы виды водорослей с различным объемом клеток, который, как известно, влияет на удельную скорость роста и приводит к существенным различиям ее значений, достигающим 2–3 раза даже в пределах одной таксономической группы (Стельмах, 2010; Finkel, 2001). Учитывая видоспецифичность максимальной скорости роста, все величины μ для каждого из трех видов диатомовых водорослей были нормированы на ее максимальное для вида значение (μмакс.). В результате между величиной µ/µмакс и относительной переменной флуоресценцией наблюдалась линейная зависимость с коэффициентом детерминации, равным 0,96, отражающая тесную связь активности реакционных центров фотосистемы 2 с удельной скоростью роста водорослей. Видно, что при значениях Fv/Fm в диапазоне от 0,15 до 0,20 µ/µмакс не превышало 0,20, то есть составляло не более 20% от максимальной величины (рис. 1б). Вероятно, недостаток биогенных веществ, и прежде всего азота, в клетках микроводорослей приводит к снижению фотохимической эффективности ФС2, а затем и к низким значениям скорости роста водорослей. Повышение относительной переменной флуоресценции до 0,30–0,40 приводило к увеличению нормированных значений удельной скорости роста до 0,30–0,55. И наконец, наиболее высоким величинам отношения Fv/Fm (0,55–0,65) соответствовали нормированные значения μ, достигавшие 100% от максимальных, что свидетельствовало об отсутствии лимитирования роста водорослей по биогенным веществам. Видно, что величина Fv/Fm при условиях, оптимальных для роста, в пределах одной таксономической группы водорослей (в данном случае диатомовых водорослей) различается между видами незначительно по сравнению с удельной скоростью роста. Как показано ранее, этот параметр очень слабо зависит также и от таксономической принадлежности водорослей (Berges et al., 1996; Johnson, 2015).
Рис.1. Связь удельной скорости роста (µ), а также ее нормированных значений (µ/µмакс.) с относительной переменной флуоресценцией (Fv/Fm) для трех видов диатомовых водорослей: S. costatum, D. brightwellii и C. socialis Fig.1. The relationship between the specific growth rate (μ) and its normalized values (μ/μmax) with the relative variable fluorescence (Fv / Fm) for three species of diatoms: S. costatum, D. brightwellii and C. socialis
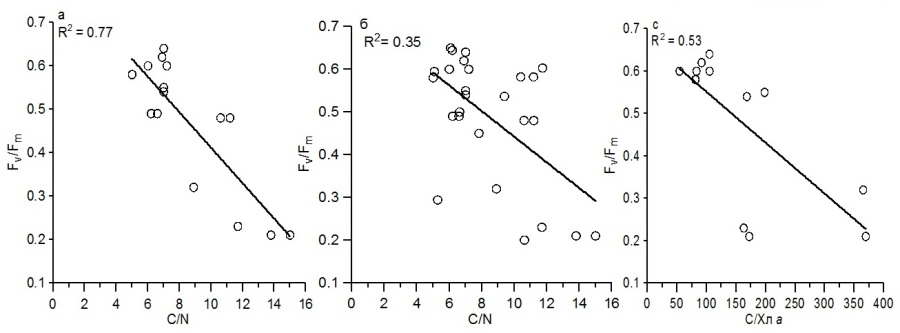
Концентрация биогенных веществ в воде определяет внутриклеточное их содержание, которое влияет на структурные и функциональные параметры водорослей. Исследования, выполненные на культуре диатомовой водоросли C. curvisetus, позволили показать, что отношение Fv/Fm связано линейной зависимостью с внутриклеточным отношением между органическим углеродом и азотом (С/N). Максимальная относительная переменная флуоресценция (0,55–0,60) наблюдалась, когда отношение С/N в клетках водорослей составляло 6–8 (рис. 2а). По мере увеличения последнего приблизительно в 2 раза относительная переменная флуоресценция снижалась до 0,20. Эти результаты подтверждены данными, полученными ранее для зеленой водоросли Dunaliella tertiolecta (Levasseur et al., 1993), диатомовых Thallassiosira weissflogii (Levasseur et al., 1993) и T. pseudonana (Parkhil et al., 2001), а также динофитовой водоросли Prorocentrum minimum (Johnson, 2015). Они свидетельствуют о важной роли биогенных веществ, и, прежде всего, азота в регуляции величины относительной переменной флуоресценции в водорослях. Что касается минерального фосфора, то в экспериментах на культуре динофитовой водоросли Amphidinium carterae выявлено, что его дефицит в среде вызывает слабое снижение отношения Fv/Fm, не превышавшее 20% от максимального значения (Li et al., 2016). Подобный эффект наблюдался у диатомовых водорослей при ограничении кремния (Hoffmann et al., 2007). Построение общей зависимости между отношением С/N и величиной относительной переменной флуоресценции для трех видов диатомовых водорослей S. costatum, D. brightwellii и C. curvisetus (рис. 2б) показало значительное ослабление связи между этими параметрами (R2 = 0,35). Вероятно, это обусловлено в основном видовыми различиями в величинах С/N отношения у исследованных водорослей. Поэтому наиболее тесные связи Fv/Fm со структурными параметрами следует ожидать для одного вида водорослей. Важный структурный показатель микроводорослей – отношение С/Хл а, достаточно быстро реагируя на изменение абиотических условий среды (света, температуры и биогенных веществ), также отражает функциональное состояние водорослей (Geider, 1987).
Рис. 2. Зависимость относительной переменной флуоресценции от внутриклеточного атомарного отношения С/N (мг-ат·мг-ат-1) для C. curvisetus (а), а также для трех видов водорослей: S. costatum, D. brightwellii и C. curvisetus (б); связь относительной переменной флуоресценции с отношением С/Хл а для C. curvisetus (c) Fig.2. Dependence of the relative variable fluorescence from the intracellular C/ N ratio (mg-atom·mg-atom-1) for C. curvisetus (a), as well as for three species of algae: S. costatum, D. brightwellii, and C. curvisetus (b); the relation of the relative variable fluorescence with the ratio C/Chl a for C. curvisetus (c)
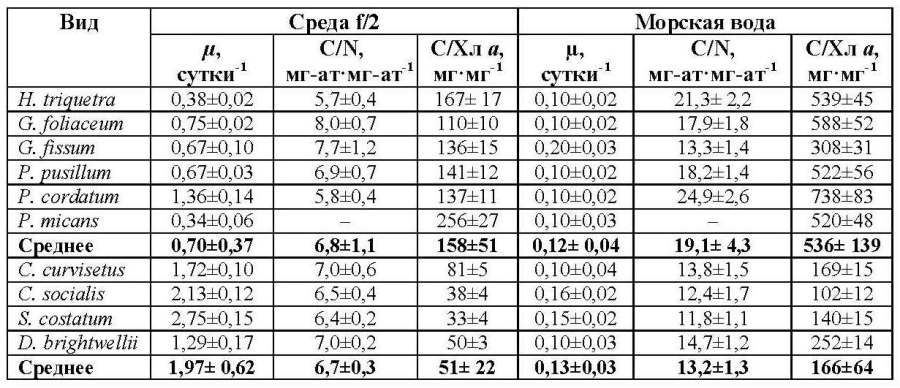
Вероятно, между этим отношением и величиной Fv/Fm при определенных условиях может наблюдаться корреляция. Так в экспериментах на культуре C. curvisetus получено, что относительная переменная флуоресценция связана обратной линейной зависимостью с величиной отношения между органическим углеродом и хлорофиллом а (рис. 2с). Это обусловлено тем, что при понижении внутриклеточного содержания биогенных элементов, и в том числе азота, в процессе роста исследуемого вида в накопительной культуре параметр Fv/Fm постепенно уменьшается. При этом происходит снижение удельного содержания хлорофилла а в расчете на единицу органического углерода, а значит С/Хл а отношение возрастает. Тогда как для трех видов диатомовых водорослей S. costatum, D. brightwellii и C. curvisetus подобная связь не выявлена, что обусловлено существенными межвидовыми различиями в величине С/Хл а. Степень видовых и таксономических различий в значениях удельной скорости роста микроводорослей, а также отношения С/Хл а и атомарного отношения между органическим углеродом и азотом (С/N) при оптимальных биогенных условиях и при биогенном лимитировании неодинакова. При достаточной обеспеченности водорослей биогенными веществами удельная скорость роста динофитовых водорослей различалась между видами приблизительно в 4 раза (табл.). Самые высокие значения (1,36 сутки-1) были выявлены для P. cordatum, а самые низкие (0,34 сутки-1) – для P. micans. Межвидовые отличия наблюдаются и по структурным параметрам водорослей. Так, отношение между органическим углеродом и хлорофиллом а различалось приблизительно в 2 раза. Наиболее низкое отношение С/Хл а (110) отмечено у G. foliaceum, тогда как наиболее высокое (256) наблюдалось у P. micans. Такой структурный параметр, как отношение С/N изменялся слабо. Представленные значения у динофитовых водорослей были в диапазоне 5,8–8,0, составив в среднем 6,8. У диатомовых водорослей при оптимальных световых и температурных условиях, а также достаточном количестве питательных веществ различия в величине удельной скорости роста не превышали 2 раза. Самые высокие значения этого параметра (2,75 сутки-1) отмечены для мелкого вида S. costatum, тогда как самые низкие (1,29 сутки-1) – для крупного вида D. brightwellii. Значения отношения С/Хл а изменялась от 33 у S. costatum до 81 у C. curvisetus. Величины отношения С/N между видами диатомовых водорослей почти не различались, а среднее значение было равно 6,7. По средним значениям исследуемых структурных и функциональных параметров для диатомовых и динофитовых водорослей хорошо видно, что удельная скорость роста диатомовых видов в 2,8 раза выше, чем у динофитовых и составила в среднем 1,97 сутки-1. Это обусловлено, прежде всего, разным относительным содержанием хлорофилла а в клетках исследуемых видов водорослей. Видно, что величина С/Хл а отношения у диатомовых водорослей составила 51, что соответствует 2% удельного содержания хлорофилла в расчете на единицу органического углерода. Тогда как у динофитовых водорослей С/Хл а отношение было в 3 раза выше и отражает достаточно низкое удельное содержание хлорофилла в клетках, составляющее в среднем 0,6% от органического углерода. Это является, как известно, основной причиной более слабого роста динофитовых водорослей по сравнению с диатомовыми (Tang, 1996). По нашим данным, средние значения отношения C/N для этих групп водорослей практически не различались.
Таблица. Удельная скорость роста (µ), отношение между органическим углеродом и азотом (C/N), а также между органическим углеродом и хлорофиллом а (C/Хл а) в культурах морских планктонных водорослей при оптимальных по биогенным веществам условиях и при их недостатке Table. The specific growth rate (μ), the ratio between organic carbon and nitrogen (C/N), and between organic carbon and chlorophyll a (C/Chl a) in marine plankton algae cultures under optimal nutrient conditions and under their limitation
Примечание: представлены средние значения параметров и стандартное отклонение
При крайнем недостатке биогенных веществ в среде различия в величинах удельной скорости роста в целом незначительны как в пределах одной таксономической группы, так и между ними. Среднее значение для диатомовых видов водорослей составило 0,13 сутки-1, а для динофитовых – 0,12 сутки-1. В тоже время межвидовые различия в величине отношения между органическим углеродом и азотом в клетках водорослей возросли. Его минимальное значение среди динофитовых водорослей (13,3) получено для G. fissum, а максимальное (24,9) – для P. cordatum , а в среднем составило 19,1. У диатомовых водорослей отношение C/N между видами различалось намного слабее, чем у динофитовых, и составило в среднем 13,2. В этих условиях отношение между органическим углеродом и хлорофиллом а у динофитовых водорослей достигло в среднем 536 при колебаниях от 308 до 738. Это свидетельствует о крайне низком удельном содержании хлорофилла в клетках водорослей, составившем 0,14–0,33% от органического углерода. У диатомовых водорослей С/Хл а отношение было в среднем в 3 раза ниже и находилось в диапазоне от 102 до 252, что соответствует 0,40–1% хлорофилла от органического углерода.
Заключение Полученные результаты и анализ данных, опубликованных в литературе, свидетельствуют о том, что относительная переменная флуоресценция хлорофилла in vivo практически не зависит от видовой и таксономической принадлежности планктонных микроводорослей. Тогда как удельная скорость их роста и отношение С/Хл а в клетках при оптимальных по биогенным веществам условиях среды подвержены значительной межвидовой и таксономической изменчивости. При высокой степени биогенного лимитирования эти различия наблюдаются только для С/Хл а отношения. В море в условиях совместного влияния факторов среды и различий в видовой и таксономической структуре фитопланктонного сообщества при оценке его функционального состояния по трем представленным выше параметрам следует учитывать видовой состав и таксономическую принадлежность водорослей.
Благодарности. Автор статьи выражает искреннюю благодарность Мансуровой И.М. и Кожемяка А.Б. за помощь в выполнении работы.
Работа выполнена в рамках государственного задания по направлению «Функциональные, метаболические и токсикологические аспекты существования гидробионтов и их популяций в биотопах с различным физико-химическим режимом».
Список литературы 1. Заика В.Е. Удельная продукция водных беспозвоночных. – Киев: Наук. думка, 1972. – 143 с. 2. Маторин Д.Н., Венедиктов П.С., Конев Ю.Н., Каземирко Ю.В., Рубин А.Б. Использование двухвспышечного импульсного погружного флуориметра для определения фотосинтетической активности природного фитопланктона // Доклады Академии Наук. 1996. Т.350, №2. С. 256–258. 3. Парсонс Т.Р., Такахаши М., Харгрей В. Биологическая океанография. – М.: Легкая и пищ. пром-сть, 1982. – С. 89–91. 4. Стельмах Л.В., Бабич И.И. Сезонные изменения отношения органического углерода к хлорофиллу «a» в фитопланктоне прибрежных вод Черного моря в районе Севастополя // Океанология. 2003. Т. 43, № 6. С. 875–884. 5. Стельмах Л.В., Куфтаркова Е.А., Акимов А.И., Бабич И.И., Кожемяка А.Б. Использование переменной флуоресценции хлорофилла in vivo для оценки функционального состояния фитопланктона // Системы контроля окружающей среды. 2010. №13. С. 263–268. 6. Финенко З.З., Стельмах Л.В., Мансурова И.М., Георгиева Е.Ю., Цилинский В.С. Сезонная динамика структурных и функциональных показателей фитопланктонного сообщества в Севастопольской бухте // Системы контроля окружающей среды. 2017. №9 (29). С. 73–82. 7. Berges J.A., Charlebois D.O., Mauzerall D.C., Falkowski P.G. Differential effects of nitrogen limitation on photosynthetic efficiency of photosystems I and II in microalgae // Plant Physiol. 1996. V.110. P. 689 –696. 8. Berges J.A., Falkowski P.G. Physiological stress and cell death in marine phytoplankton: Induction of proteases in response to nitrogen or light limitation // Limnol. Oceanogr. 1998. V. 43. P. 129–135. 9. Finkel Z.V. Light absorption and size scaling of light-limited metabolism in marine diatoms // Limnol. Oceanogr. 2001. V.46. P. 86–94. 10. Geider R.J. Light and temperature dependence of the carbon to chlorophyll ratio in microalgae and cyanobacteria: implications for physiology and growth of phytoplankton // New Phytol. 1987. V.106. P. 1–34. 11. Grasshoff K., Ehrhardt M., Kremling K. Methods of seawater analysis. – Weinheim, Germany: Verlag Chemie, 1983. – 419 p. 12. Guillard R.R.L., Ryther J.H. Studies of marine planktonic diatoms: I. Cyclotella nana Hustedt, and Detonula confervacea (Cleve) Gran // Can. J. Microbiol. 1962. V.8, №2. P. 229–239. 13. Hoffmann L.J., Peeken I., Lochte K. Effects of iron on the elemental stoichiometry during EIFEX and in the diatoms Fragilariopsis kerguelensis and Chaetoceros dichaeta // Biogeosciences. 2007. V.4. P. 569–579. 14. Holm-Hansen O., Lorenzen C. J., Holmes R.W. Fluorometric determination of chlorophyll // J. Cons. int. Explor. Mer. 1965. V.30. P. 3–15. 15. Johnson M.D. Inducible mixotrophy in the dinoflagellate Prorocentrum minimum // J. Eukaryotic Microbiology. 2015. V.62. P. 431 – 443. 16. Kolber Z., Falkowski P.G. Use of active fluorescence to estimate phytoplankton photosynthesis in situ // Limnol. Oceanogr. 1993. V.38. P. 1646–1665. 17. Levasseur M., Thompson P. A., Harrison P.J. Physiological acclimation of marine phytoplankton to different nitrogen sources // J. Phycol. 1993. V.29. P. 587–595. 18. Li M., Shi X., Guo C., Lin S. Phosphorus deficiency inhibits cell division but not growth in the dinoflagellate Amphidinium carterae // Front. Microbiol. 2016. V.7. P. 826. doi: 10.3389/fmicb.2016.00826 19. Parkhil J.P., Maillet G., Cullen J.J. Fluorescence-based maximal quantum yield for PSII as a diagnostic of nutrient stress // J. Phycol. 2001. V.37. P. 517–529. 20. Tang E.P.Y. Why do dinoflagellates have lower growth rates // J. Phycol. 1996. V.32. P. 80–84. Статья поступила в редакцию 7.12.2017
Functional status and some structural characteristics of marine plankton algae at different levels of nutrients availability Lyudmyla V. Stelmakh Kovalevsky Institute of Marine Biological Research of RAS (Sevastopol, Russia) The influence of the nutrients content in the medium on the intracellular C/N ratio variability, the specific growth rate in cultures, the relative variable fluorescence of chlorophyll (Fv/Fm) and the ratio of C/Chl a was studied on 10 species of the Black Sea microalgae. When these parameters are changed, the closest correlation between them is observed within one species of algae, and it weakens or is absent for different species. Key words: microalgae; specific growth rate; relative variable fluorescence; intracellular nitrogen content; the ratio between organic carbon and chlorophyll a.
Об авторе Стельмах Людмила Васильевна – Stelmakh Lyudmyla V. кандидат биологических наук lustelm@mail.ru Корреспондентский адрес: 299011 г. Севастополь, пр. Нахимова, 2, Институт биологии южных морей им. А.О. Ковалевского; тел.: +7 (8692) 54-41-10.
ССЫЛКА НА СТАТЬЮ: Стельмах Л.В. Функциональное состояние и некоторые структурные характеристики морских планктонных водорослей при разном уровне обеспеченности биогенными веществами // Вопросы современной альгологии. 2018. № 1 (16). URL: http://algology.ru/1252 Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно. При перепечатке ссылка на сайт обязательна
К разделу ОБЗОРЫ, СТАТЬИ И КРАТКИЕ СООБЩЕНИЯ

|
|||
|
| ||