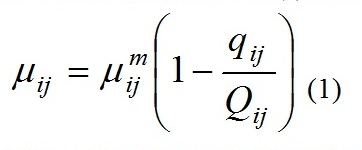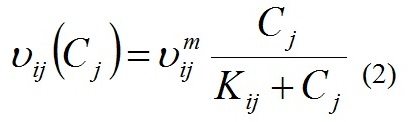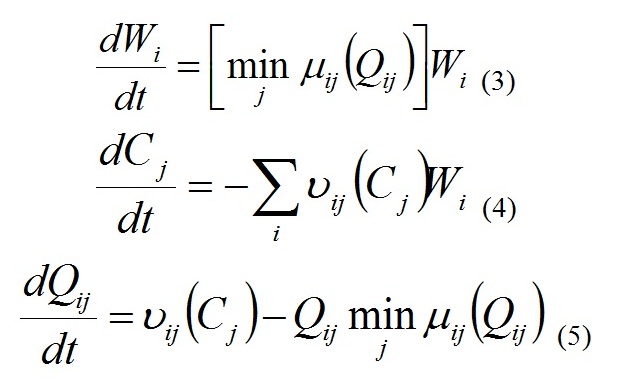|
|
|
Реакция фитопланктонного сообщества на катастрофическое наводнение (июль 2012 г.)
Паутова Л.А.1, Силкин В.А.2, Часовников В.К.2 Larisa A. Pautova, Vladimir A. Silkin, Valeriy K. Chasovnikov
1Институт океанологии им. П.П. Ширшова РАН (Москва, Россия)
УДК 574.52
Катастрофические ливни в районе г. Геленджика, произошедшие в июле 2012 г., привели к аномальному паводку и залповому выбросу в прибрежную экосистему большого количества пресной воды с высоким содержанием азота, фосфора и кремния. Концентрация этих элементов в Голубой бухте, куда впадает река Ашамба, повысилась на 2 порядка для фосфора и кремния и в 50 раз для азота. Эффект уменьшения солености на разрезе от устья р. Ашамба к центру моря прослеживался по всему шельфу вплоть до склона. Концентрация элементов питания на разрезе снижалась по экспоненциальному закону. Сообщество фитопланктона в момент паводка находилось в состоянии перехода от доминирования кокколитофорид к доминированию крупноклеточных диатомей. Залповое повышение концентрации элементов питания не привело к существенной перестройке структуры фитопланктона, реакция сообщества заключалась в существенном повышении численности и биомассы существующих доминант. Так, численность кокколитофориды Emiliania huxleyi достигала 9 млн. клеток в литре, а биомасса крупноклеточной диатомеи Pseudosolenia calcar-avis – 40,4 г/м3. При этом первые превалировали на нижних горизонтах, а вторые – на верхних. Через 10 дней после паводка эффект краткосрочного эвтрофирования сохранялся только в Голубой бухте, а через 20 дней экосистема пришла в своему обычному для этого времени года состоянию. Существенное влияние на количественные показатели фитопланктона оказывал эффект разбавления водами открытого моря. Ключевые слова: фитопланктон; паводок; диатомовые водоросли; кокколитофорида Emiliania huxleyi; Черное море.
Введение Прибрежные экосистемы функционируют в условиях высоких градиентов гидрофизических и гидрохимических параметров. Фитопланктон реагирует на изменения этих параметров, что сказывается на его структурно-функциональных характеристиках. В олиготрофных и мезотрофных сообществах определяющим является потоки биогенных элементов, от соотношения которых зависит видовая структура фитопланктона. В целом это достаточно консервативная для каждого сезона структура, которая зависит от синоптических и климатических факторов. Они могут смещать сроки появления доминант или даже привести к существенному изменению видового состава вплоть до появления новых доминант. Фитопланктонное сообщество претерпевает регулярные изменения видового состава в течение годового цикла, называемые сукцессией. Но существуют явления, которые носят экстремальный характер, и они могут привести к существенным сдвигам в экосистеме. К таким явлениям относятся ливни и сопровождающие их наводнения. Одним из таких экстремальных событий является катастрофический ливень, прошедший в районе г. Геленджик 6–7 июля 2012 г. По данным Росгидромета за вечер 6 июля в Геленджике выпало до 231 мм осадков, тем самым месячная норма осадков только за один день была превышена в 4–5 раз. За сутки с 07 часов 6 июля по 07 часов 7 июля эта величина составила 311 мм. Уровень в створе реки Ашамба (500 м от устья) поднимался на 4–6 метров, что вызвало локальное наводнение. В русле реки сформировался необычайно мощный поток (скорость 5–7 м/с), который на своем пути имел катастрофические последствия – были разрушены дома, дороги, произошла деформация русла реки и ее поймы (Куклев и др., 2014). Этот грязекаменный поток вынес в Голубую бухту крупные деревья, автомобили, постройки. Огромный объем грязекаменного материала изменил морфоструктурный облик донной поверхности бухты. По подсчетам с водосбора реки в бухту попало порядка 17,5 млн. м3 пресной воды. По другим оценкам этот показатель составлял 13 млн. м3 (Крыленко и др., 2014). Влияние повышенных осадков в речном водосборе исследуются в основном на примере экосистем эстуариев, бухт и проливов (Sylvan et al., 2007; Carstensen et al., 2015; Balch et al., 2012). Главным образом описываются изменения в абиотической части экосистемы (концентрации азота, фосфора, солености, pH и т.д.). Кроме того, экосистема Черного моря имеет свои особенности, что необходимо учитывать. Возникает вопрос, к каким последствиям приводит данное экстремальное явление на уровне биотической составляющей прибрежной экосистемы и какие перестройки в ней происходят на уровне структуры сообществ? Кроме того, интересен вопрос о пространственно-временном распространении сдвигов структурно-функциональной организации сообществ. Цель работы заключалась в исследовании реакции фитопланктонного сообщества на изменение нагрузки на прибрежную экосистему в результате экстремального природного явления – наводнения. При этом решались следующие задачи:
Материалы и методы Исследования гидрофизических, гидрохимических параметров воды, а также структурных и количественных показателей фитопланктона были проведены с борта МНИС «Ашамба» 11.07., 18.07. и 26.07. 2012 г. Пробы отбирались на стандартном разрезе от устья р. Ашамба к центру моря на станциях с глубиной 10, 20, 30 ,50, 100 и 500 м и с горизонтов от 0 до 50 м. Для гидрофизических измерений использовался CTD зонд «Sea Bird» (Sea-Bird Electronics, Inc), установленный на погружаемом приборе типа «Rozette». Морская вода на гидрохимические параметры и фитопланктон отбиралась 5-литровым батометром Нискина. Пробы фитопланктона фиксировались на борту судна 40% формалином, нейтрализованным бораксом (Na2BO3), до конечной концентрации 1% и концентрировались седиментационным методом. Идентификацию видов и подсчет числа клеток проводили с помощью светового микроскопа в счетных камерах Науманна (объем 1 мл) для микрфитопланктона и камерах Нажотта объемом 0,05 мл для пико- и нанофитопланктона. Использовали световой микроскоп (Ergaval; Karl Zeiss, Jena) с увеличением 16×20 и 16×40. Биомассу рассчитывали, приравнивая клетки по конфигурации к различным геометрическим фигурам (Hillebrand et al., 1999). В вопросах номенклатуры руководствовались современными источниками (Identifying Marine Phytoplankton, 1997; Throndsen et al., 2003, www.algaebase.org и www.marinespecies.org). Все остальные процедуры были стандартными в соответствии с общепринятыми методами (Киселев, 1969; Moncheva, Parr, 2010). Анализы на фосфаты, нитраты, нитриты, аммоний и кремний были проведены с использованием Techicon ll анализатора (Современные методы гидрохимических…, 1992; Grashoff et al., 1999). Для оценки потенциальных возможностей получения биомассы фитопланктона в случае отсутствия разбавления был проведен расчет кривой накопления по аналогии с накопительной культурой с использованием ранее апробированной модели (Silkin et al., 2013, 2016). При построении математической модели применялась концепции внутриклеточной регуляции роста водорослей и применение принципа минимума. В данном случае удельная скорость роста определяется внутриклеточным содержанием элемента, лимитирующего рост в соответствии с моделью Друпа (Droop, 1974):
где: μij и μijm – соответственно удельная и максимальная удельная скорости роста i-го вида на j-ом элементе питания; Qij и qij – соответственно текущее и минимальное содержание j-го элемента питания в биомассе i-го вида. Удельная скорость поглощения элемента питания определяется его концентрацией в среде, следуя модели Михаэлиса-Ментен:
где: υij и υijm – соответственно удельная и максимальная удельная скорости поглощения j -го элемента питания i -ым видом; Kij – константа полунасыщения для j-го элемента питания у i-го вида.
Функции Wi и Cj обозначают концентрации биомассы i-го вида и j-го элемента питания соответственно. Проводили расчеты для описания динамики биомассы Pseudosolenia calcar-avis с момента начала паводка, константы уравнений брали из работы (Silkin et al., 2013).
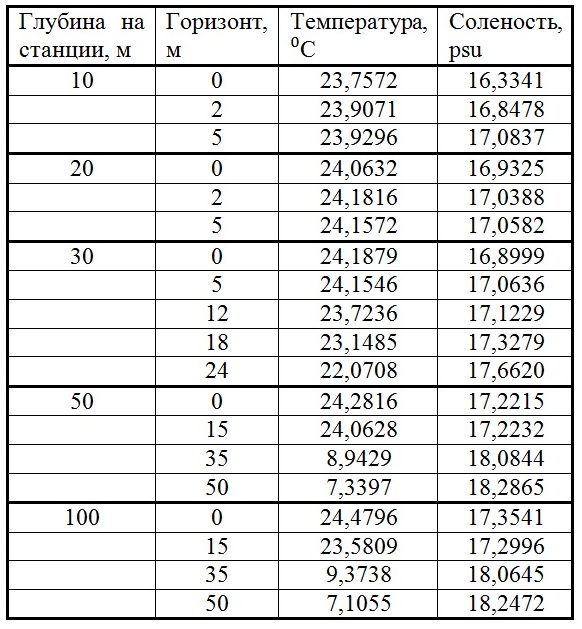
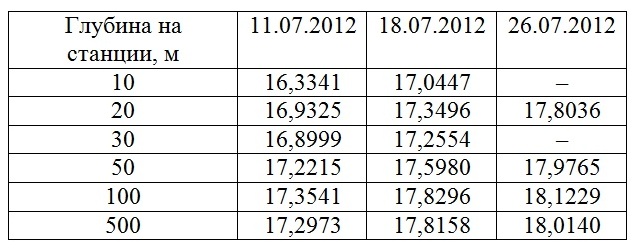
Результаты Гидрофизические показатели После катастрофического ливня верхний перемешанный слой (ВПС) 11.07.2012 г. достигал глубины 20 м (табл. 1). На станциях с глубиной 10, 20 и 30 м влияние пресных вод прослеживается во всем слое ВПС, однако наибольшее распреснение регистрировалось в верхнем пятиметровом слое воды. Распространение повышенного (залпового) речного стока от устья к центру моря можно проследить по изменению солености и температуры (пресные воды более холодные) в поверхностном слое воды (табл. 2). Если 11.07 пресные воды растекались по поверхности до наиболее удаленной станции с глубиной 500 м, то к 18.07 такое влияние распространялось до станции с глубиной 100 м, а 26.07 небольшое распреснение фиксировалось только в Голубой бухте.
Таблица 1. Изменение температуры и солености по вертикали
Таблица 2. Изменение солености в поверхностном слое воды (psu) на различных станциях шельфа 11.07, 18.07 и 26.07.2012 г.
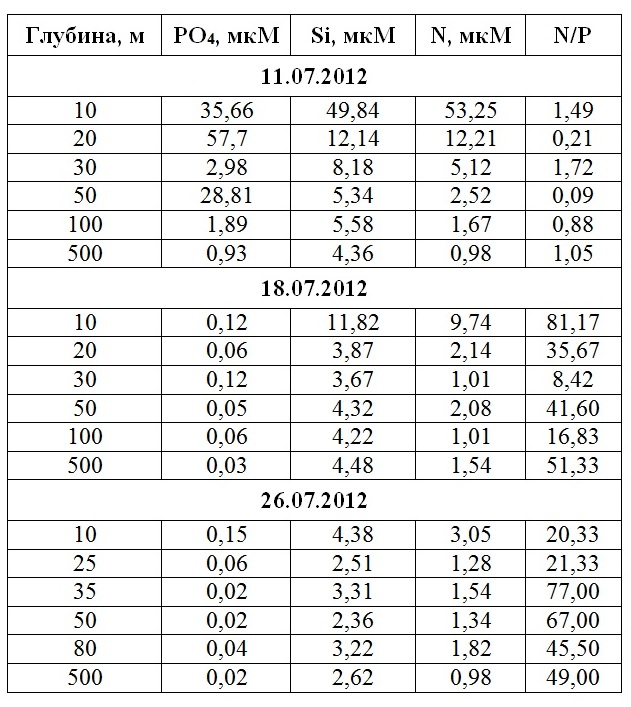
Гидрохимические условия Повышенный сброс пресной воды оказал существенное влияние на концентрацию азота, фосфора и кремния (табл. 3). Так, 11.07.2012 г. концентрация фосфора и кремния на станции с глубиной 10 м (Голубая бухта) превысила среднемноголетние величины на 2 порядка, а концентрация азота – в 50 раз. При этом регистрировался существенный градиент элементов минерального питания по вертикали, так в Голубой бухте (станция с глубиной 10 м) концентрация азота на поверхности моря многократно превышала концентрацию этого элемента на глубине 5 м (53,2 против 3,22 мкМ).
Таблица 3. Концентрация азота, фосфора и кремния (мкМ) в поверхностном слое воды на различных станциях разреза от устья р. Ашамба к центру моря 11.07, 18.07 и 26.07. 2012 г.
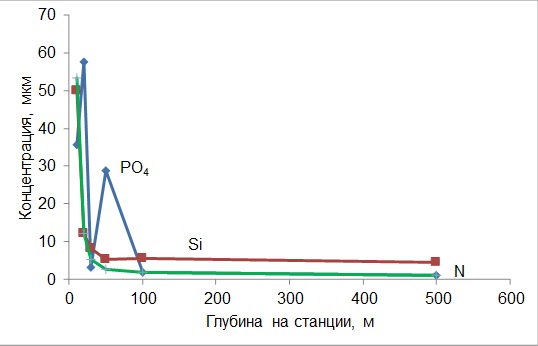
Что касается изменения концентрации по разрезу от устья к центру моря, то падение концентрации азота и кремния носит экспоненциальный характер (рис. 1). Такой закономерности не подчиняется изменение концентрации фосфора – здесь зарегистрировано 2 максимума на станциях с глубинами 20 и 50 м. Уже к 18.07.2012 г. повышенные концентрации элементов питания прослеживаются только в Голубой бухте, а к 26.07.2012 г. экосистема приходит к состоянию, где гидрохимический режим не отличается от средне многолетнего (табл. 3). Соотношение азота к фосфору во время паводка близко к 1, т.е. намного меньше соотношения Редфилда. Уже к 18.07.2012 г. этот показатель возвращается к обычным для этого времени значениям.
Рис. 1. Изменение концентрации минеральных форм азота (нитраты+нитриты+аммоний),
Реакция фитопланктонного сообщества на катастрофический паводок Основу фитопланктонного сообщества составляют две таксономические группы: диатомовые и кокколитофориды (табл. 4). При этом основной доминантой в диатомовой компоненте была крупная водоросль Pseudosolenia calcar-avis, вклад которой в биомассу диатомей составлял свыше 90%. Кокколитофориды были представлены только одним видом – Emiliania huxleyi. Данный вид 11.07.2012 г. На фоне высоких концентраций азота и фосфора формировал «цветение» в ВПС, максимальная численность 9,6×106 кл/л была зарегистрирована на горизонте 15 м станции с глубиной 20 м. В верхних горизонтах от 0 до 5 м наблюдалось «цветение» крупноклеточной диатомеи P. calcar-avis. Так, на горизонте 5 м станции с глубиной 20 м была зафиксирована биомасса 40,4 г/м3. Биомасса такого уровня не регистрировалась за весь период многолетних исследований. Также аномально высокая биомасса 28,5 г/м3 была обнаружена на горизонте 25 м станции с глубиной 500 м.
[при нажатии на ссылку открывается полный текст таблицы]
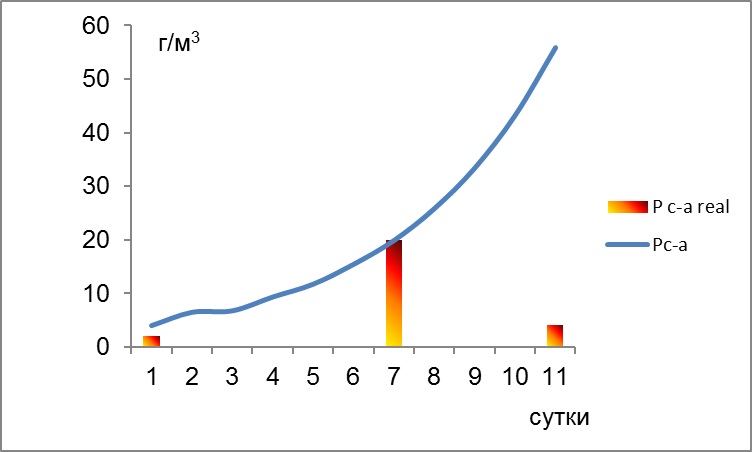
18.07.2012 г. «цветение» кокколитофориды продолжилось, хотя численные показатели были значительно ниже. Если на станциях с глубинами 10 и 20 м «цветение» регистрировалось во всем столбе воды, то на более дальних от устья станциях данный вид интенсивно вегетировал на нижних горизонтах. Диатомея Pseudosolenia calcar-avis демонстрировала более высокий уровень биомассы на близких к устью станциях. Очевидно, что кроме поглощения азота, фосфора и кремния доминатами фитопланктона, основную роль в снижении концентрации элементов играет разбавление за счет перемешивания с водой открытого моря. Для оценки максимального уровня биомассы фитопланктона на существующих концентрациях элементов минерального питания рассчитывали кривую накопления для станции над глубиной 50 м. Данный расчет показал, что в случае отсутствия разбавления на 11 день после паводка, т.е. к 18.07.2012 г., биомасса диатомеи Pseudosolenia calcar-avis могла достигнуть 50 г/м3 (рис. 2).
Рис. 2. Динамика реальной биомассы (столбики) диатомеи Pseudosolenia calcar-avis и теоретическая линия, рассчитанная по модели (уравнение 3, см. раздел Материал и методы).
26.07.2012 г. влияние наводнения на структуру фитопланктона практически не прослеживается, на всех станциях она типична для этого сезона.
Обсуждение В результате многолетних исследований нами выделены основные доминанты фитопланктона северо-восточной части Черного моря, выявлены характерные особенности годовой динамики, основанной на смене доминант (Pautova et al., 2007). Показано, что в весенний период доминантами чаще всего становятся мелкоклеточные диатомеи из рода Pseudo-nitzschia, в конце весны и начале лета практически ежегодно доминируют кокколитофориды (Emiliania huxleyi), а летом доминантами становятся крупноклеточные диатомеи Pseudosolenia calcar-avis и Proboscia alata (Silkin et al., 2013). Кокколитофориды доминируют при относительно низких концентрациях азота и относительно высоких концентрациях фосфора, при этом соотношение азота к фосфору значительно ниже соотношения Редфилда (Silkin et al., 2014). Переход от доминирования кокколитофорид к доминированию крупноклеточных диатомей P. calcar-avis и P. alata происходит обычно во второй половине июня. Однако в 2012 г. в первой половине июня было зарегистрировано необычайно мощное «цветение» кокколитофорид, оно продолжалось и во второй половине июня. Съемки, проведенные 22.06.2012 г., показали, что кокколитофориды продолжали доминировать и на шельфе наблюдалось «цветение» этой водоросли. Максимальная численность вида превышала 3 млн. кл/л. Но уже в начале июля в сообществе стали занимать лидирующие по биомассе позиции крупноклеточные диатомеи, а именно Pseudosolenia calcar-avis. К началу паводка отмечалась переходная стадия от кокколитофорид к крупноклеточным диатомеям. Смена доминант всегда связана с изменением гидрохимического режима вод. При доминировании крупных диатомей концентрация фосфора очень низка, а соотношение азота к фосфору очень велико и значительно превышает соотношение Редфилда. Пришедшие с паводком воды значительно повысили концентрацию элементов питания. При этом соотношение азота к фосфору значительно ниже соотношения Редфилда. Это стимулировало развитие кокколитофорид, для которых такое соотношение оптимально, и наблюдалось «цветение» этой водоросли с количественными показателями, более характерным и даже превышающим таковые в конце весны и начале лета, т.е. периода доминирования кокколитофорид. Другими словами, добавление порции элементов питания как бы вернуло сообщество к прошлой стадии. В то же время повышенные концентрации элементов и, прежде всего кремния, стимулировало развитие крупноклеточной диатомеи P. calcar-avis. Надо отметить, что в период доминирования крупноклеточных диатомовых, обычно происходящего летом, концентрация кремния существенно снижается за счет потребления, и он может стать лимитирующим рост элементом. Это связано с тем, что существующие концентрации кремния могут быть сравнимы с константой полунасыщения для поглощения кремния этим видом (Krause et al., 2012). Особенность реакции сообщества на паводковые воды и связанным с этим залповым повышением концентрации элементов питания в том, что не происходит смена доминант, структура сообщества остается прежней, а существенно повышаются количественные показатели. При этом происходит разделение видов по вертикали: крупноклеточная диатомея превалирует в верхних горизонтах, а кокколитофорида в нижних. Исключение составляет зарегистрированная высокая биомасса Pseudosolenia calcar-avis на горизонте 25 м станции с глубиной 500 м. Такое явление, скорее всего, связано с существованием вихрей в период их затухания, когда поверхностные воды опускаются вниз. Неизменность структуры сообщества при однократном введении большого количества элементов питания связана с коротким временем самого импульса. Расчеты показали, что существенную роль в формировании количественных показателей фитопланктона играет эффект разбавления. Действительно, если бы не существовало смешивания прибрежных вод и вод открытого моря, биомасса фитопланктона была бы такой высокой, что не исключена возможность возникновения заморных явлений при разложении этой биомассы.
Исследование проводилось в рамках государственного задания ФАНО (тема № 0149-2018-0009).
Список литературы
Статья поступила в редакцию 14.06.2018.
The response of the phytoplankton community to a catastrophic flood (July 2012) Larisa A. Pautova1, Vladimir A. Silkin2, Valeriy K. Chasovnikovv2 1Shirshov Institute of Oceanology RAS (Moscow, Russia) Catastrophic showers in the Gelendzhik area, which occurred in July 2012, led to an abnormal flood and instantaneous emission to the coastal ecosystem of a large amount of fresh water with a high content of nitrogen, phosphorus and silicon. The concentration of these elements in the Blue Bay, where the river Ashamba flows, increased by 2 orders of magnitude for phosphorus and silicon and 50 times for nitrogen. The effect of a decrease in salinity on a section from the mouth of the river Ashamba to the center of the sea was traced all over the shelf up to the slope. The concentration of the nutrients on the section decreased exponentially. The phytoplankton community was in a state of transition from the dominance of coccolithophorids to the dominance of large-cell diatoms at the time of the flood. An increase of the nutrients concentration did not lead to a significant restructuring of the phytoplankton structure, the community's reaction consisted in a substantial increase in the number and biomass of existing dominants. Thus, the number of coccolithophorides Emiliania huxleyi reached 9 million cells per liter, and the biomass of the large cell diatom Pseudosolenia calcar-avis was 40.4 g/m3. The former prevailed on the lower horizons, while the latter prevailed on the upper horizons. Ten days after the flood, the effect of short-term eutrophication persisted only in the Blue Bay, and after 20 days the ecosystem came to its usual state for this time of year. A significant effect on the quantitative indicators of phytoplankton is the effect of dilution of the coastal water by the waters of the open seas. Key words: phytoplankton; flood; diatoms; coccolithophore Emilianua huxleyi; Black Sea.
Об авторах Паутова Лариса Альбертовна − Pautova Larisa A. кандидат биологических наук
Силкин Владимир Арсентьевич - Silkin Vladimir A. доктор биологических наук
Часовников Валерий Кузьмич – Chasovnikov Valeriy K. кандидат географических наук
Корреспондентский адрес: Россия, 353470, Краснодарский край, г. Геленджик, ул. Просторная 1-г. Телефон/факс 8-861-41-280-89.
ССЫЛКА НА СТАТЬЮ: Паутова Л.А., Силкин В.А., Часовников В.К. Реакция фитопланктонного сообщества на катастрофическое наводнение (июль 2012 г.) // Вопросы современной альгологии. 2018. № 2 (17). URL: http://algology.ru/1295
При перепечатке ссылка на сайт обязательна
На ГЛАВНУЮ
|
|||
|
| ||