|
|
|
|
по Материалам Международной конференции «Экологическая физиология водных фототрофов: распространение, запасы, химический состав и использование» VII Сабининские чтения 1 ноября - 25 декабря 2018 г. Механизмы регуляции числа клеток в клеточной цепи диатомей
Федоров А.В., Лифанчук А.В. Aleksey V. Fedorov, Anna V. Lifanchuk
Институт океанологии им. П.П. Ширшова РАН, Южное отделение (Геленджик, Россия)
УДК 574.52
Предложена гипотеза, описывающая механизм формирования и распада цепочек клеток, по которой количество клеток в цепи определяется разницей скоростей деления клеток и разделения цепочек на отдельные клетки, а максимальное число клеток в цепочке достигается при наибольшей скорости деления. Показано, что добавка фосфора стимулирует образование более длинных цепочек. Разработана простая математическая модель динамики числа клеток в клеточных цепях. Ключевые слова: культура фитопланктона; колонии диатомовых водорослей; удельная скорость роста; скорость разделения цепочек.
Введение Одной из важных задач экологии является адекватная оценка текущего состояния экосистемы, а так как морской фитопланктон играет важнейшую роль в формировании экосистем морей и океанов и составляет основу пищевой цепи (Falkowski et al. 1998; Field et al., 1998), оценка его состояния становится первоочередной задачей. Фитопланктон определяет функционирование морской экосистемы, и без оперативной оценки его состояния невозможен прогноз развития экосистемы в будущем. Исходя из вышесказанного, создание инструментария оперативной оценки функциональных свойств фитопланктона представляется актуальной задачей. Несмотря на большое таксономическое разнообразие сообществ фитопланктона, именно диатомовые водоросли создают основную продукцию в море и являются основой биологической помпы (Smetachek, 1999). Как один из возможных методов оперативной оценки предлагается использовать длину клеточных цепей диатомовых в качестве показателя состояния популяции (Mikaelyan, 1995; Lifanchuk, 2014). Характерным свойством пелагических диатомей является формирование колоний при помощи различных типов соединений (Суханова, 1981), что, с одной стороны, в значительной мере способствует их плавучести, а с другой – препятствует выеданию зоопланктоном (Kiørboe, Saiz, 1995; Ward et al., 2012). Наличие в экосистеме хищника с избирательным предпочтением к размерам жертвы может также изменить длину цепочек (Peter, Sommer, 2012). Среднее число клеток в клеточной цепи у различных видов и популяций диатомей может значительно варьировать, что зависит от таксономического положения, условий среды и физиологического состояния этих популяций (Прошкина-Лавренко, 1955). Существуют различные предположения о причинах преобладания длинных или, наоборот, коротких цепочек клеток диатомовых. При этом колониям клеток отводится различная функциональная роль вплоть до сопротивления выеданию зоопланктоном (Reynolds, 1988; Long et al., 2007; Selander et al., 2011; Bergkvist et al., 2012; Bjærke et al., 2015). Образующие цепочки клеток диатомовые водоросли воспринимают химический сигнал копепод и реагируют на него путем изменения длины цепи. При этом снижается их выедание более чем в 10 раз (Bergkvist et al., 2012). Другие исследователи считают, что факторы среды влияют не только на морфологию фитопланктона, но и на изменчивость среднего числа клеток в цепи (Kling, 1993). Существуют данные экспериментов, в которых показано, что количество клеток в цепочке Skeletonema costatum повышалось при увеличении концентрации элементов минерального питания и температуры (Takabayashi et al., 2006; Ebrahimi, Salarzadeh, 2016), а в других исследованиях – с увеличением солености (Balzano et al., 2011; Ebrahimi, Salarzadeh, 2016). Некоторые авторы считают, что удлинение клеточной цепи и увеличение объемов клеток связано с перемешиванием водным масс. Это объясняется большим поступлением питательных веществ в клетку (Mukherjee et al., 2015). Интуитивно ясно, что длина цепи отражает физиологическое состояние популяции, и используя этот показатель, можно создать новый инструмент анализа продукционных свойств и прогноза их развития в будущем. Однако механизм образования и распада цепочек не выяснен, и факторы, влияющие на среднее количество клеток в цепочке диатомей, остаются малоизученными. В настоящей работе выдвигается гипотеза, объясняющая формирование цепочек колониальных диатомовых водорослей. В основу её положен следующий механизм: процесс образования цепочек клеток связан с процессом деления клеток и их разделения. Прирост числа клеток в цепи зависит от скорости роста биомассы колониальных диатомовых водорослей, которая, в свою очередь, определяется оптимальным для каждого вида набором фактором среды (свет, концентрация и соотношение элементов минерального питания) (Lifanchuk, 2014; Silkin et al., 2014). Разделение цепочек на отдельные клетки не связано с ростом биомассы образующих колонии диатомей, а контролируется физическими свойствами среды, такими как температура и турбулентность (Lindenmayer, 1968). Целью работы было проверить данную гипотезу на непротиворечивость экспериментальным исследованиям и результатам математического моделирования.
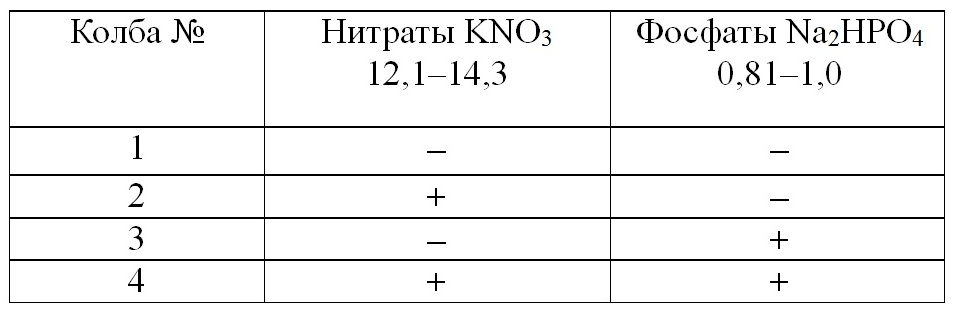
Материалы и методы В период 2014–2018 гг. в береговой лаборатории Южного отделения Института океанологии РАН (г. Геленджик) проводили эксперименты с накопительной культурой фитопланктона, в которых исследовали влияние добавок элементов минерального питания (нитратов и фосфатов) на структуру фитопланктонного сообщества. Объектом исследования служила смешанная культура водорослей (кокколитофорид, диатомовых и динофитовых), полученная на основе природного сообщества фитопланктона. Пробы были собраны с борта малого научно-исследовательского судна «Ашамба». Отбор проб производился с горизонта 0 м на станции стандартного разреза от Голубой бухты к центру моря. Горизонт выбран в связи с тем, что в поверхностном слое присутствуют виды, характерные для всего слоя, а концентрации биогенных элементов близки нулю или на порядки ниже, чем в нашем опыте сразу после произведения добавки (Часовников и др., 2018), что способствует чистоте эксперимента. Станция располагалась над глубиной 50 м (район срединного шельфа). Морскую воду отбирали пластиковой пятилитровой емкостью и разливали по бутылям с одновременной фильтрацией через два слоя мельничного газа № 36 для удаления зоопланктона. Выращивание проводили в колбах Эрленмейера емкостью 500 мл, где объем культуральной среды составлял 200 мл. В эксперименте использовали аппарат для культивирования (термолюминостат), где температура среды соответствовала температуре на поверхности моря на момент отбора проб. Интенсивность падающего света поддерживали на уровне 58–61 мкмоль/м2 ФАР. Идентификацию видов (www.algaebase.org; www.marinespecies.org; Tomas, 1997) и подсчет числа клеток проводили ежедневно на световом микроскопе в счетной камере Ножотта объемом 0,05 мл. Биомассу рассчитывали методом «истинного объема» (Киселев, 1969), при этом использовались данные собственных измерений. Основным объектом исследования в данной работе была природная популяция фитопланктона, выращиваемая в культуре. Изучалась реакция фитопланктона на изменение параметров среды, которые наиболее значимы в исследуемый период, с применением хорошо разработанных методов планирования эксперимента (Максимов, Федоров, 1969; Федоров, Кустенко, 1972). Для экспериментов использовалась схема полного факторного эксперимента (Максимов, 1980). В Табл. 1 знак «-» указывает, что добавление элементов минерального питания не производили, а «+» означает, что добавление было выполнено. В качестве факторов среды нами была выбрана концентрация азота и фосфора. Это связано с тем, что район северо-восточной части Черного моря относится к мезотрофному типу (Паутова и др., 2007) и, следовательно, существенное значение для продуктивности экосистемы имеют элементы минерального питания. Концентрация кремния не рассматривалась как гипотетический регулятор, поскольку она находится в прибрежных водах на достаточно высоком уровне (Yakushev et al., 2007). Основными видами диатомей, образующими клеточные цепи, за период исследования были Chaetoceros socialis var. autumnalis Prosckina-Lavrenko, Leptocylindrus danicus P.T. Cleve, Skeletonema costatum (Greville) P.T. Cleve. В меньшем количестве в экспериментах росли Thalassionema nitzschioides (Grunow, 1862) Van Heurck, Pseudo-nitzschia pseudodelicatissima (Hasle) Hasle, Chaetoceros curvisetus P.T. Cleve и Dactyliosolen fragilissimus (Bergon) G.R. Hasle. Эти водоросли входили в лидирующий комплекс видов северо-восточной части Черного моря в различные сезоны и годы (Паутова и др., 2007; Силкин и др., 2013). В экспериментах применяли периодический (накопительный) режим культивирования (Силкин, Хайлов, 1988). Исходными параметрами служили биомасса фитопланктона в стационарной фазе накопительной культуры, максимальное и среднее число клеток в цепи доминирующих видов. Добавку элементов минерального питания (нитратов в форме KNO3 и фосфатов в форме Na2HPO4) производили в начале эксперимента по единой схеме, представленной в табл. 1. Закономерности роста и динамики среднего числа клеток в клеточной цепи проверяли на соответствие экспериментальным данным методами математического моделирования в вычислительном эксперименте с использованием уравнений кинетики роста биомассы от концентраций элементов минерального питания (Силкин и др., 2013).
Таблица 1. План экспериментов по влиянию добавок элементов минерального питания на продукционные свойства (мкМ/л) Table 1. The plan of experiments on the effect of additives of mineral nutrition elements on the production properties (μM /L)
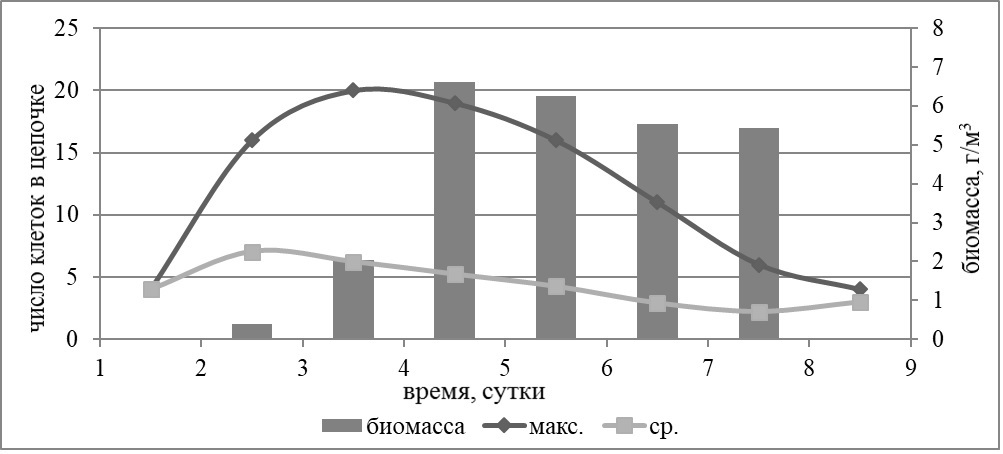
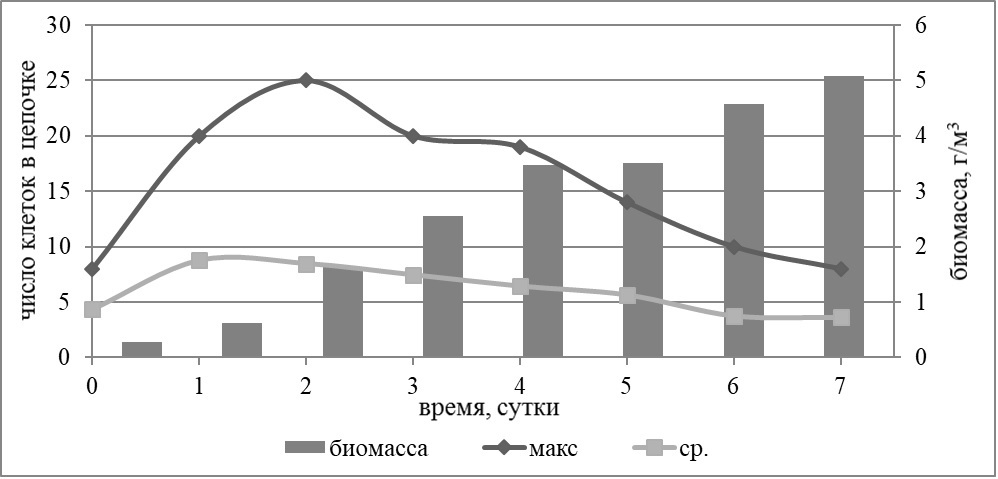
Результаты Как показали результаты многолетних полевых исследований, структура фитопланктонного сообщества северо-восточной части Черного моря чаще всего определяется двумя таксономическими группами – кокколитофоридами и диатомеями, и реже динофлагеллятами. В наших экспериментах динофлагелляты не давали значимого прироста биомассы, реагировали на добавки элементов минерального питания диатомеи и кокколитофориды. Доминирование одной из групп зависит от соотношения концентраций азота и фосфора (Silkin et al., 2014). Кокколитофориды доминируют при низких концентрациях азота и высоких концентрациях фосфора, т.е. при низких соотношениях азота к фосфору (ниже соотношения Редфилда). Для доминирования диатомей необходимо, чтобы концентрации азота и фосфора были высокими, а их соотношение близко к соотношению Редфилда. В опыте №1 (дата постановки 28 мая 2014 г., длительность 9 суток) в вариантах с добавкой фосфора доминировала кокколитофорида Emiliania huxleyi. В вариантах с добавкой азота, а также совместной добавкой азота и фосфора доминантой являлась диатомовая водоросль Skeletonema costatum. В варианте без добавок преобладала диатомея Dactyliosolen fragilissimus. Максимальный прирост биомассы для всех видов водорослей происходил в варианте с совместной добавкой нитратов и фосфатов. Наибольшей биомассы (6,61 г/м3) достигла мелкоклеточная диатомея S. costatum. Максимальное число клеток в колонии данной диатомеи было достигнуто на третий день эксперимента и составляло 20 клеток в первой повторности опыта. Динамика максимального и среднего числа клеток в цепочке представлена на рис. 1. Наибольшее среднее число клеток в цепи (7 клеток) зарегистрировано на второй день опыта.
Рис. 1. Динамика биомассы и максимальной (макс.) и средней (ср.) длины клеточной цепи Skeletonema costatum в опыте №1 с совместной добавкой азота и фосфора Fig. 1. Dynamics of biomass and maximum (макс.) and average (ср.) length of the Skeletonema costatum cell chain in the experiment №1 with simultaneous addition of nitrogen and phosphorus
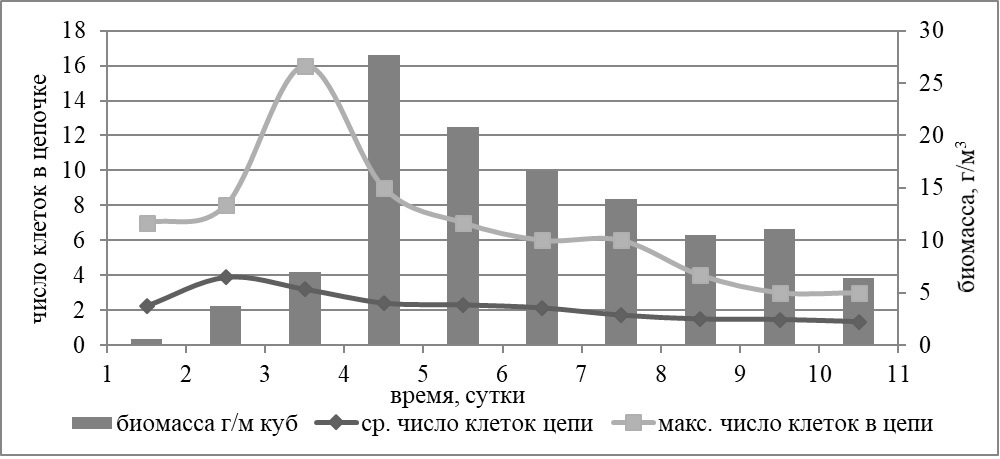
Во второй повторности опыта с совместной добавкой азота и фосфора наибольшее среднее число клеток в цепи отмечено на третий день эксперимента – 6 клеток в цепочке. Максимальная число клеток в цепи – 16. Общая закономерность, заключающаяся в том, что цепочки содержат большее число клеток в первые дни эксперимента при максимальной нелимитированной скорости роста, подтверждается результатами анализа обеих повторностей опыта. В варианте опыта без добавок элементов минерального питания четкой зависимости числа клеток в цепи Skeletonema costatum от роста биомассы не выявлено. Как и во всех остальных вариантах опыта, без добавок повторяется тенденция к снижению среднего числа клеток в цепочке к концу эксперимента. В колбах с добавкой только нитратов эта тенденция наиболее сильно выражена. Среднее число клеток в цепи в начале эксперимента составляло 5–6 клеток, а к концу эксперимента 1–2 клетки. Цепочка S. costatum максимальной длины во всём эксперименте зарегистрирована в варианте опыта с добавкой фосфора, и она состояла из 24 клеток в одной из повторностей. В опыте №2 (дата постановки 2 июня 2015 г., длительность 11 суток) во всех вариантах, исключая добавку фосфатов, доминировала диатомея Leptocylindrus danicus. В варианте с добавкой фосфора доминантой являлась кокколитофорида Emiliania huxleyi. Максимальный прирост биомассы для всех видов водорослей был отмечен в варианте с совместной добавкой нитратов и фосфатов. В этом эксперименте практически во всех вариантах опыта доминировала колониальная мелкоклеточная диатомея Leptocylindrus danicus. В варианте с совместной добавкой азота и фосфора биомасса этой диатомеи достигала 27 г/м3 на четвертый день эксперимента, тогда как биомассы остальных водорослей в культуре были на порядок ниже. Наибольшее максимальное число клеток в цепочках (16) зарегистрировано на третий день опыта, а среднее (4 клетки) – на второй день.
Рис. 2. Динамика биомассы и максимальной и средней длины клеточной цепи Leptocylindrus danicus в опыте №2 с совместной добавкой азота и фосфора Fig. 2. Dynamics of biomass and maximum and average length of the cell chain of Leptocylindrus danicus in the experiment №2 with the simultaneous addition of nitrogen and phosphorus
После достижения стационарной фазы рост Leptocylindrus danicus прекратился, а цепочки постепенно разделялись. К концу эксперимента встречались преимущественно одиночные клетки. В контрольном варианте и с добавкой фосфора четкой картины динамики не выявлено. Конкуренцию Leptocylindrus danicus в этом опыте составляла также колониальная мелкоклеточная диатомея Skeletonema costatum. Она росла при совместной добавке азота и фосфора, приблизившись к Leptocylindrus danicus лишь в одной повторности (4.2) с биомассой 11,8 г/м3. В остальных вариантах опыта биомасса S. costatum была на порядок ниже биомассы конкурирующего с ней L. danicus и не отвечала ростом на добавки азота и фосфора по отдельности. Такая же картина наблюдалась и в варианте без добавок. В опыте №3, поставленном в марте 2016 г. и длившемся 7 суток, сообщество фитопланктона было представлено в основном мелкоклеточными колониальными диатомеями весеннего комплекса Skeletonema costatum и Chaetoceros socialis, максимальные биомассы которых достигали 5 г/м3 (в опыте с совместной добавкой азота и фосфора), и кокколитофоридой Emiliania huxleyi. В варианте без добавок элементов минерального питания лидировала по биомассе диатомея Chaetoceros socialis (1004,9 мг/м3). С совместной добавкой азота и фосфора происходил интенсивный рост биомассы обоих видов диатомовых водорослей. Максимально длинная зарегистрированная цепочка S. costatum в этом эксперименте состояла из 25 клеток и отмечена на второй день эксперимента. Наибольшее же среднее число клеток в цепи (9 клеток) посчитано на 2-й день опыта. Максимальное число клеток в клеточной цепи оставалось высоким (более 20 клеток в цепочке) в течение четырех дней. Динамика биомассы и числа клеток в цепи S. costatum показана на рис. 3.
Рис. 3. Динамика биомассы и максимальной и средней длины клеточной цепи Skeletonema costatum в опыте №3 с совместной добавкой азота и фосфора Fig. 3. Dynamics of biomass and maximum and average length of the cell chain of Skeletonema costatum in the experiment №3 with the simultaneous addition of nitrogen and phosphorus
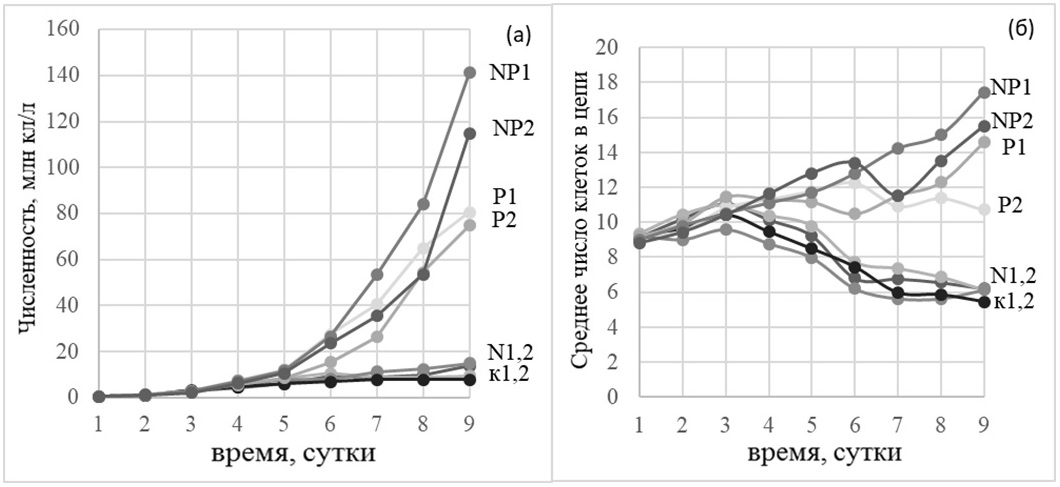
В вариантах опыта с добавками азота и фосфора по отдельности и без добавок совсем биомасса S. costatum оказалась на порядок ниже, ее рост был прерывистым, а получившиеся графики плохо отражали тренды динамики размеров колоний. Добавка фосфора способствовала образованию водорослью S. costatum длинных цепочек (максимальное число клеток – 21 на 5-й и 6-й день эксперимента, среднее 8–11 клеток). В данном опыте диатомея Chaetoceros socialis была конкурентом Skeletonema costatum. Максимальная биомасса при совместной добавке элементов минерального питания составляла 4,87 г/м3. Рост Chaetoceros socialis продолжался до конца опыта. Максимальная удельная скорость роста в обеих повторностях составляла около 1 сут-1. Максимальная длина клеточной цепи достигала 19 клеток на четвертые сутки. В обеих повторностях наибольшее среднее число клеток в клеточной цепи наблюдалось на второй день эксперимента (около 8 клеток). К концу эксперимента средняя длина клеточной цепочки составляла 2 клетки. Динамика этого показателя повторяет таковую в других опытах, для вариантов с четко выраженным ростом биомассы. В вариантах без добавок элементов минерального питания, с добавкой только азота или только фосфора биомасса не превышала 1 г/м3. В связи с небольшим ростом и конкуренцией за элементы минерального питания с Skeletonema costatum, рост Chaetoceros socialis был неравномерным, а динамика числа клеток в цепочке неявной, без четких трендов роста и пиков. Таким образом, добавка азота не влияла на увеличение длины клеточной цепи. В колбах с добавлением фосфора биомасса (0,8 г/м3) не превышала значений таковой для контрольного варианта эксперимента без добавок. Однако в обеих повторностях среднее число клеток в цепи на протяжении всего эксперимента было выше, чем в варианте с добавками азота и в варианте без добавок. В феврале 2018 г. в смешанной культуре на основе природного сообщества фитопланктона северо-восточной части Черного моря преобладала диатомовая водоросль Skeletonema costatum. В опыте №4 (дата постановки 28 февраля 2018 г., продолжительность 9 суток) на культуру, как и в предыдущих опытах, воздействовали путем добавок в начале эксперимента нитратов и фосфатов по отдельности и совместно в соотношении 14:1. Однако для того, чтобы показать зависимость среднего числа клеток в цепи S. costatum от скорости роста, в данном опыте вместо биомассы использовали численность. Это обусловлено необходимостью снижения разброса значений при подсчёте клеток. Так как значение биомассы критически зависит от случайного распределения размеров клеток по выборке, а численность более равномерно возрастает при росте культуры, именно последняя использовалась для расчёта скоростей роста. Во всех предыдущих опытах разброс значений рассчитанной скорости роста не позволил достоверно оценить взаимосвязь этого параметра со средним числом клеток в цепи. Во всех повторностях опыта доминирующим видом была колониальная диатомея S. costatum. Доля других видов составляла не более 10%. Это в первую очередь связано с тем, что исходная природная культура взята в прибрежной области Черного моря, где данная диатомея встречается регулярно. Кроме того, в зимний период вода обогащена элементами минерального питания, но при условиях короткого светового дня, относительно низкой интенсивности света и довольно сильного перемешивания S. costatum – один из немногих видов водорослей, способных давать большую численность в данный период. Это обуславливается высокой удельной скоростью роста данной диатомеи при достаточно эффективном фотосинтетическом аппарате (более того, дальнейшие опыты показали угнетающую роль интенсивного освещения на скорость роста Skeletonema costatum). Лимитирующий фактор определяет потенциал роста фитопланктона, и в феврале в опытах с добавками биогенных элементов таким фактором была концентрация фосфора, т.к. именно на его добавку отреагировал фитопланктон. С другой стороны, достижение стационарной фазы в данном опыте значительно отставало от экспериментов в более поздние сезоны, что можно объяснить меньшей длительностью освещения культуры (короткий световой день в феврале). Со свето-темновым периодом, характерным для мая–июня стационарная фаза достигалась уже после четвертых суток. Таким образом, можно предположить, что световые условия наравне с концентрациями биогенов являются определяющими факторами, регулирующими скорость роста. Численность Skeletonema costatum в вариантах опыта с добавкой фосфора к концу эксперимента была практически на порядок выше, чем в вариантах без добавки или с добавкой только азота, что говорит об отсутствии лимитирования по азоту в поверхностном слое моря и недостатке минерального фосфора. Максимальная численность клеток отмечена в конце эксперимента в варианте с совместной добавкой азота и фосфора и составила около 140 млн кл/л. На число клеток в цепочках также оказала влияние добавка фосфора. Это показывают кривые динамики численности и числа клеток в цепочках S. costatum на рис. 4.
Рис. 4. Динамика численности (а) и среднего числа клеток в цепи (б) Skeletonema costatum в эксперименте №4 с добавками азота и фосфора; к – контроль (без добавок), N – добавка азота, Р – добавка фосфора, цифрами – номер повторности. Fig. 4. The dynamics of cell number (a) and the average number of cells in the chain (b) of Skeletonema costatum in an experiment №4 with addition of nitrogen and phosphorus on February 28, 2018;
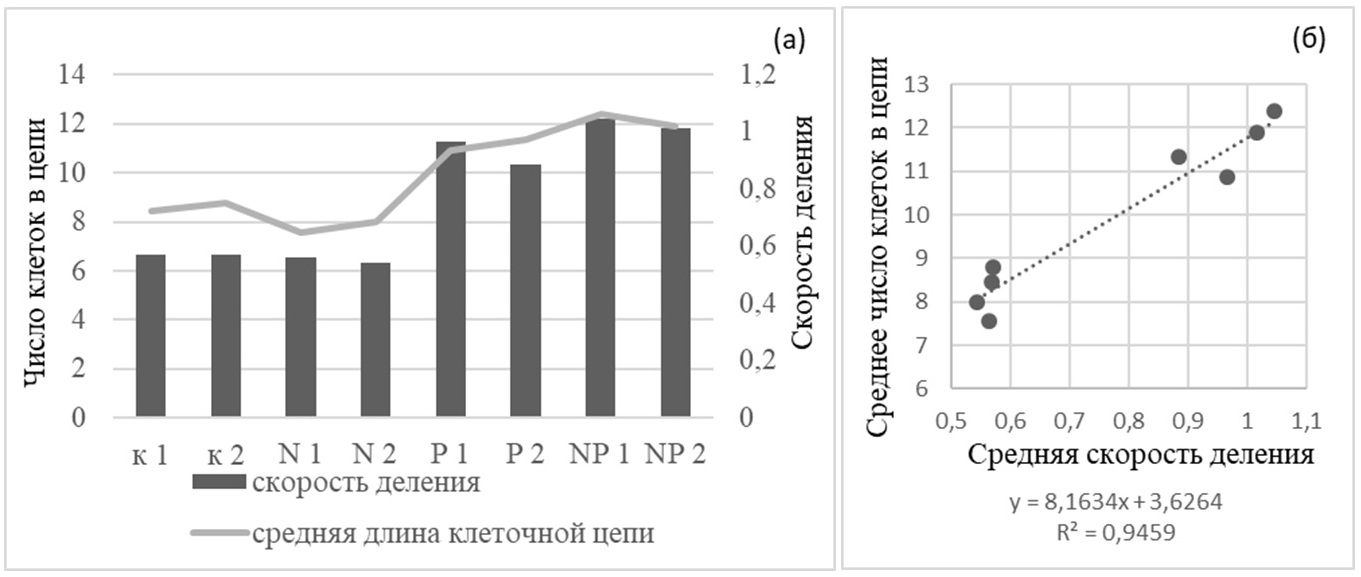
Среднее число клеток в цепи и усредненная за весь период эксперимента скорость деления S. costatum хорошо коррелируют (рис. 5).
Рис. 5. Диаграмма значений среднего числа клеток в цепи и скорости деления клеток Skeletonema costatum (а) и зависимость этих двух параметров друг от друга (б) в эксперименте №4 с добавками азота и фосфора; к – контроль (без добавок), N – добавка азота, Р – добавка фосфора, цифрами – номер повторности. Fig. 5. Diagram of average values of number of cells in the chain compared with growth rate of Skeletonema costatum (a) and correlation of these two parameters on each other (b) in the experiment №4 with addition of nitrogen and phosphorus; к – control (without additives), N – addition of nitrogen, P – addition of phosphorus, numeric – number of repetition.
Данные эксперимента не противоречат выдвинутой гипотезе, однако в данном случае есть сложность с разделением роли факторов – с одной стороны скорость роста очевидно регулирует число клеток в цепочке Skeletonema costatum. С другой стороны, как показали и некоторые предыдущие опыты, цепочки клеток становились длиннее при добавке фосфора и без значимого увеличения скорости роста. Несмотря на очевидную зависимость скорости роста и числа клеток в клеточных цепях Sk. сostatum в данном опыте от концентрации фосфора (рис. 5), его специфическое влияние на число клеток в цепочках диатомей требует осторожности при интерпретации результатов и дальнейших исследований. Также значительную роль в формировании или распаде колоний играют физические факторы среды: свет, температура и перемешивание. Интенсивность света, например, является главным регулятором скорости роста при отсутствии лимитирования элементами минерального питания. Таким образом, описанные выше результаты проведенных экспериментов (кривые динамики биомассы и числа клеток в цепи, а также зависимость от скорости роста, показанная в опыте № 4) позволяют утверждать, что предложенная нами гипотеза механизма образования и распада цепочек клеток имеет право на существование. Число клеток в клеточной цепи, согласно положенной в основу данной работы гипотезы, изменяется вслед за изменением удельной скорости роста в ходе эксперимента. Скорость роста зависит от условий среды, а скорость разделения более консервативна, но также может зависеть от таких факторов, как температура, интенсивность света и др. (в нашей модели данный параметр рассматривается как постоянная). Далее предложено математическое выражение обнаруженной закономерности формирования и разделения цепочек и проверено на простейшей математической модели. В практике исследований по непрерывному культивированию микроорганизмов широкое применение нашла модель Моно (Monod, 1950), связывающая удельную скорость роста и концентрацию вещества – источника углерода и энергии С: где µ − удельная скорость роста, сут-1; Для описания роста биомассы и поглощения элементов минерального питания использовали следующие дифференциальные уравнения:
где W – биомасса, мг/м3; Для выражения зависимости удельной скорости роста от концентрации элемента питания использовали формулу (1) Моно (Monod 1950). Количество клеток в цепочке, согласно нашей гипотезе, принималось пропорциональным разности текущей удельной скорости роста и скорости разделения клеток, и для выражения этих процессов использовалось следующее выражение: где N – число клеток в клеточной цепи; Для построения численной модели роста биомассы фитопланктона и поглощения двух основных лимитирующих элементов в водной среде – азота и фосфора, использовали следующие уравнения в численном виде:
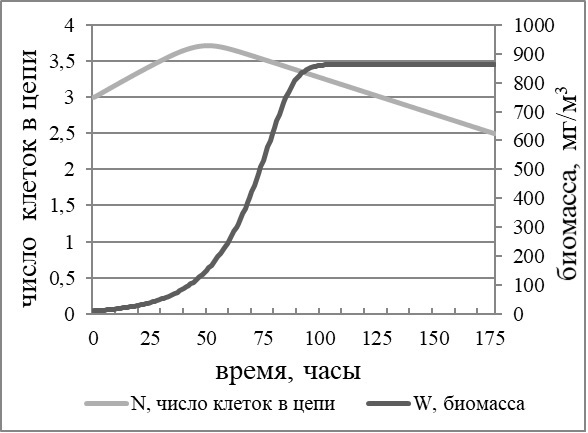
где Wn – биомасса в момент времени n, мг/м3; В качестве начальных условий вычислительного эксперимента брали биомассу и начальные концентрации N и P, соотвествующие их значениям в реальных экспериментах (N – 12–14 мкМ; P – 0,8–1 мкМ). Значения константы полунасыщения и максимальной удельной скорости роста, а также содержания выбранного элемента в клетке брали из литературных данных для мелкоклеточных колониальных диатомей. Так как эти константы сильно зависят от других условий в опыте (сильнее всего от света), то для общего описания закономерности роста вводили значения этих параметров, смоделированные кривые которых максимально близки динамике роста культур в наших экспериментах. Значения скорости распада клеточных цепей g и введенного коэффициента а подбирали для соответствия численной модели данным собственных опытов. В результате моделирования получается экспоненциальная кривая биомассы с насыщением. В процессе роста идет поглощение элементов минерального питания. Наибольшая биомасса достигается на четвертые-пятые сутки (100 часов). Кривая динамики числа клеток в цепи (рис. 6), описанная численной моделью, повторяет таковые, полученные в результате анализа экспериментальных исследований. Максимальная длина цепочки наблюдается в период экспоненциальный фазы роста и достигается на вторые сутки (50 часов) после начала эксперимента (время наступления стационарной фазы роста, как и наибольшего числа клеток в цепи, зависит от начальных условий эксперимента). После исчерпания азота и фосфора в растворенном виде (по результатам моделирования согласно кинетике поглощения), скорость распада клеточных цепей превышает удельную скорость роста, и к концу опыта среднее число клеток в цепочке составляет 1–2 клетки.
Рис. 6. Зависимость длины клеточной цепи от роста биомассы в вычислительном эксперименте Fig. 6. Dependence of number of cells in cell chain on growth of biomass in a calculative experiment
В природных популяциях на момент постановки экспериментов среднее число клеток в цепочках основных видов диатомовых было невелико. Помещение культур микроводорослей в благоприятные условия и добавки элементов минерального питания способствовали увеличению удельной скорости роста. Максимальные длины цепочек наблюдались в период экспоненциального роста. При отсутствии лимитирования элементами минерального питания, что происходило в вариантах опыта с совместной добавкой азота и фосфора, уже в первые сутки эксперимента фитопланктон делится с удельной скоростью роста, близкой к максимальной. От одного до трех дней, пока еще элементов минерального питания либо в среде, либо во внутриклеточных запасах хватает для поддержания экспоненциального роста (согласно моделям кинетики роста и поглощения элементов минерального питания), длится период нарастания колоний. Со временем растворенных в воде нитратов и фосфатов вследствие активного поглощения этих элементов минерального питания фитопланктоном становится всё меньше, что приводит к снижению темпов деления. Таким образом, анализ результатов проведенных опытов демонстрирует непротиворечивость выдвинутой нами гипотезы экспериментальным данным. Все виды показали наибольшее среднее число клеток в клеточной цепи в фазу максимальной скорости роста биомассы фитопланктона. Это говорит о том, что у всех видов, как у пеннатных Pseudo-nitzschia pseudodelicatissima и Thalassionema nitzschioides (Lifanchuk, 2014), так и у центрических Leptocylindrus danicus, Skeletonema costatum и представителей рода Chaetoceros, независимо от размера и формы, а также типов соединения клеток, существует единая закономерность динамики образования клеточных цепочек. Исходя из полученных результатов, можно утверждать, что выявленная нами закономерность образования и распада цепочек действительно существует, и данный параметр можно использовать при оценке состояния популяций мелкоклеточных колониальных диатомей.
Авторы заявляют об отсутствии конфликта интересов, требующего раскрытия в данной статье. Исследование проведено в рамках темы госзадания № 0149-2019-0014 ИО РАН.
Список литературы
Статья поступила в редакцию 15.12.2018
Об авторах Фёдоров Алексей Владимирович − Fedorov Aleksey V. младший научный сотрудник, Институт океанологии им.П.П.Ширшова РАН, Южное отделение, Геленджик, Россия (P.P. Shirshov Institute of Oceanology RAS, Southern branch, Russia, Gelendzhik)
Лифанчук Анна Викторовна − Lifanchuk Anna V. кандидат биологических наук
Корреспондентский адрес: Россия, 353470, Краснодарский край, г. Геленджик, ул. Просторная 1-г. Телефон/факс 8-861-41-280-89.
ССЫЛКА НА СТАТЬЮ: Федоров А.В., Лифанчук А.В. Механизмы регуляции числа клеток в клеточной цепи диатомей // Вопросы современной альгологии. 2019. № 1 (19). С. 8–22. URL: http://algology.ru/1429 DOI - https://doi.org/10.33624/2311-0147-2019-1(19)-8-22 Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно. При перепечатке ссылка на сайт обязательна
Mechanisms of regulation of the number of cells in the cell chain of diatoms Aleksey V. Fedorov, Anna V. Lifanchuk Shirshov Institute of Oceanology RAS, Southern Branch (Gelendzhik, Russia) We proposed hypothesis that describes the mechanism of formation and disintegration of cell chains, according to which the number of cells in a chain is determined by the difference in the rates of cell division and the separation of chains into individual cells, and the maximum number of cells in the chain is achieved at the highest division rate. Experiments showed that the addition of phosphorus stimulates the formation of longer chains. We developed a simple mathematical model of the dynamics of the number of cells in cell chains. Key words: phytoplankton culture; colonies of diatoms; specific growth rate; rate of chains separation.
Fedorov A.V., Lifanchuk A.V. Mechanisms of regulation of the number of cells in the cell chain of diatoms. Voprosy sovremennoi algologii (Issues of modern algology). 2019. № 1 (19). P. 8–22. URL: http://algology.ru/1429 DOI - https://doi.org/10.33624/2311-0147-2019-1(19)-8-22
Authors Fedorov Aleksey V. ORCID - https://orcid.org/0000-0003-4673-6649 Shirshov Institute of Oceanology RAS, Southern Branch, Russia, Gelendzhik
Lifanchuk Anna V. ORCID - https://orcid.org/0000-0001-9953-7374 Shirshov Institute of Oceanology RAS, Southern Branch, Russia, Gelendzhik
References
Dear colleagues! If you want to receive the version of the article in PDF format, write to the editor,please and we send it to you with pleasure for free. When reprinting a link to the site is required
К разделу ОБЗОРЫ, СТАТЬИ И КРАТКИЕ СООБЩЕНИЯ

|
|||
|
| ||
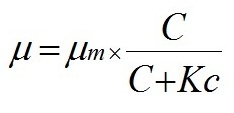
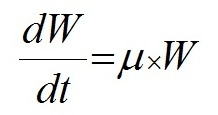
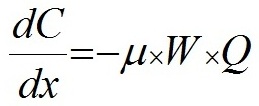
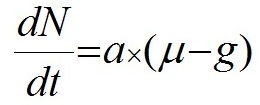
.jpg)