|
|
|
Диатомовые водоросли и их использование в стратиграфических и палеогеографических исследованиях
Стрельникова Н.И.1, Гладенков А.Ю.2 Nina I. Strelnikova, A.Yu. Gladenkov
1Санкт-Петербургский государственный университет (Санкт-Петербург, Россия)
УДК 582.26
Приводится обзор современных сведений о диатомовых водорослях – одноклеточных микроорганизмах с кремнистым панцирем, относящихся к классу Bacillariophyceae. Дана краткая характеристика биологии, морфологии, строения панциря и его структурных элементов, принципов систематики диатомей, а также биогеографии и экологии. Обсуждаются роль диатомовых в осадконакоплении и особенности сохранности их остатков в осадочных отложениях. Особое внимание уделено вопросам, связанным с эффективным использованием данных по ископаемым диатомовым водорослям при детальном биостратиграфическом расчленении, датировании и корреляции древних осадочных толщ (прежде всего, морских), а также проведении реконструкций условий окружающей среды прошлого. Дана общая характеристика зональных шкал по диатомеям, разработанных для расчленения морских отложений мела и кайнозоя. Обсуждаются основные черты эволюции диатомей. Ключевые слова: диатомовые водоросли; биология; морфология; строение панциря; экология; биогеография; биогенное кремненакопление; использование в биостратиграфии и реконструкциях условий окружающей среды прошлого; основные черты эволюции.
Вместо предисловия. Несколько лет назад у наших коллег из Института нефтегазовой геологии и геофизики им. А.А. Трофимука Сибирского отделения РАН, которые одновременно являются преподавателями в Новосибирском государственном университете, возникла идея создания учебника или учебного пособия по палеоальгологии. В первую очередь, в него планировалось включить разделы по различным группам микроводорослей (прежде всего, динофлагеллатам, диатомовым, силикофлагеллатам, кокколитофоридам), так как изучение их остатков с успехом используется при дробном биостратиграфичеком расчленении, датировании и корреляции отложений мезозоя и кайнозоя, а также проведении реконструкций параметров окружающей среды прошлых эпох. Предполагалось, что подобная сводка будет полезна как для студентов, изучающих основы палеонтологии, стратиграфии и палеоэкологии, так и для их преподавателей. В частности, профессора Н.И. Стрельникову и меня попросили написать обзорную главу по диатомовым водорослям, что мы и сделали. Однако в силу разных причин осуществить публикацию задуманного издания не удалось. В настоящее время Н.И. Стрельникова, которая является одним из наиболее известных и авторитетных российских специалистов в области изучения ископаемых диатомей, отошла от активной научной и педагогической деятельности. Однако не хотелось, чтобы работа, в которую она вложила свой труд, осталась необнародованной. Тем более, как мне кажется, написанный нами обзор во многом не утерял своей актуальности. Ознакомление с ним может представлять интерес для специалистов (прежде всего, молодого поколения) различных отраслей биологии, геологии и географии. В этой связи, заручившись согласием соавтора, я взял на себя смелость предложить нашу рукопись (с некоторыми исправлениями и дополнениями) для публикации в качестве статьи в журнал «Вопросы современной альгологии». А.Ю. Гладенков, доктор геолого-минералогических наук
1. Введение 1.1. По современным представлениям диатомовые водоросли – обширный класс Bacillariophyceae отдела Охрофитовых (Ochrophyta) водорослей, относящихся к царству Страминопилы (Stramenopiles) (Adl et al., 2005). Для Охрофитовых водорослей характерны ультраструктурные признаки: обычно два жгутика неравной длины и разного строения. Более длинный жгутик покрыт двумя рядами трехчастных мастигонем, второй — короткий, гладкий, с базальным вздутием. В переходной зоне имеется переходная спираль. Оболочка хлоропластов состоит из 4 мембран: 2 собственно мембраны хлоропласта и 2 мембраны хлоропластной эндоплазматической сети. Если хлоропласт расположен около ядра, то в большинстве случаев наружная мембрана хлоропластной эндоплазматической сети продолжается в виде наружной мембраны ядра. Ламеллы хлоропластов трехтилакоидные с опоясывающей ламеллой. Вся хлоропластная ДНК собрана в форме кольцеобразного нуклеоида; митохондрии с трубчатыми кристами (Белякова и др., 2006; Ботаника …, 2007). Сходное положение диатомей в системе органического мира принято в учебнике Algae (Van den Hoek et al., 1995). Однако это не единственное представление о положении диатомовых водорослей в системе органического мира. Во многих изданиях диатомовые рассматриваются в ранге отдела Bacillariophyta c тремя классами: Coscinodiscophyceae, Bacillariophyceae, Fragilariophyceae (Round et al., 1990). На эту сводку ориентируется большинство диатомологов. В отечественных изданиях было принято выделять в составе отдела Bacillariophyta два класса: Centrophyceae и Pennatophyceae (Диатомовые водоросли СССР ..., 1988). 1.2. Первые изображения диатомей датируются 1703 г., когда Антони ван Левенгук усовершенствовал световой микроскоп, увеличивающий в 300 раз. Он обратил внимание на странную коричневатую ладьевидную форму, ползавшую в препарате, и назвал ее «овсяным животным». Такое название было вызвано представлениями о делении органического мира, существовавшего со времен Аристотеля: весь органический мир подразделялся на животных, обладавших способностью двигаться, и неподвижные растения. С тех пор интерес к этой группе организмов привлек многих исследователей, которые изучали ныне живущих диатомей. В конце XVIII – начале XIX века шло накопление представлений и предварительных данных о диатомеях. В 1797–1802 гг. Г. Шантранс (Girod Chantrans) описывает разные формы, дискутируя, животные ли это, растения, или органокристаллы. В 1805 г. швейцарский ботаник О.П. Декандоль (Augustin Pyramus De Candolle) установил род Diatoma, от которого произошло название всей группы «диатомовые» (от греческого «разделенные пополам»). Более научное направление в изучении диатомовых начинается с 1817 г. и связано с именем К.Л. Нитсша (Christian Ludwig Nitzsch, 1817). Он соединил все диатомовые, известные в то время, в один род Bacillaria, главным признаком которого являлась призматическая, палочковидная форма. К.Л. Нитсш описал деление этих организмов на две части и выявил неразрушаемость их оболочек после отмирания. Неподвижные формы он относил к растениям, а подвижные рассматривал как животных. Название отдела Bacillariophyta дано по этому типовому роду. В 1834 г. немецкий ботаник Ф. Кютцинг (Friedrich Traugott Kützing) установил химический состав оболочек диатомовых, доказав, что они кремнеземные. Первые сведения об ископаемых диатомовых водорослях появились в середине XIX века, а начало их интенсивного изучения относится ко второй половине XX века. «Остатки» диатомовых описывались многими натуралистами. Среди пионеров таких исследований были Х.Г. Эренберг (Christian Gottfried Ehrenberg) и Д.У. Бейли (Jacob Whitman Bailey), первые работы которых по ископаемым диатомеям появились в 1836–1838 гг. В 1854 г. опубликован атлас Microgeologie, в котором Х.Г. Эренберг привел рисунки и описания разных групп ископаемых организмов, в том числе диатомей. Большое внимание исследователей, в частности, привлекали залежи белых легких пород, которые, как показали исследования, состояли из створок диатомей и получили название «organisches Polirschifers», «Kieselerde», «инфузорная земля», «кизельгур» и «диатомит». В начале XIX в. начинаются масштабные исследования Мирового океана. В 1849 г. с помощью прибора, сконструированного американцем Д.М. Бруком, с океанического дна были отобраны первые образцы грунта. Пробы (желто-коричневая глина), взятые с борта шхуны «Дельфин», были отправлены профессору Д.У. Бейли, который в 1838 г. начал изучение ископаемых диатомей из района Уэст Пойнта (West Point) в США. В России первая работа по диатомовым опубликована в 1854 г. Ф.И. Вейссе (Johann Friedrich von Weisse), а в 1873 г. Илья Григорьевич Борщов опубликовал на русском языке первую обзорную работу, в которой привел все известные к этому времени сведения о диатомеях.
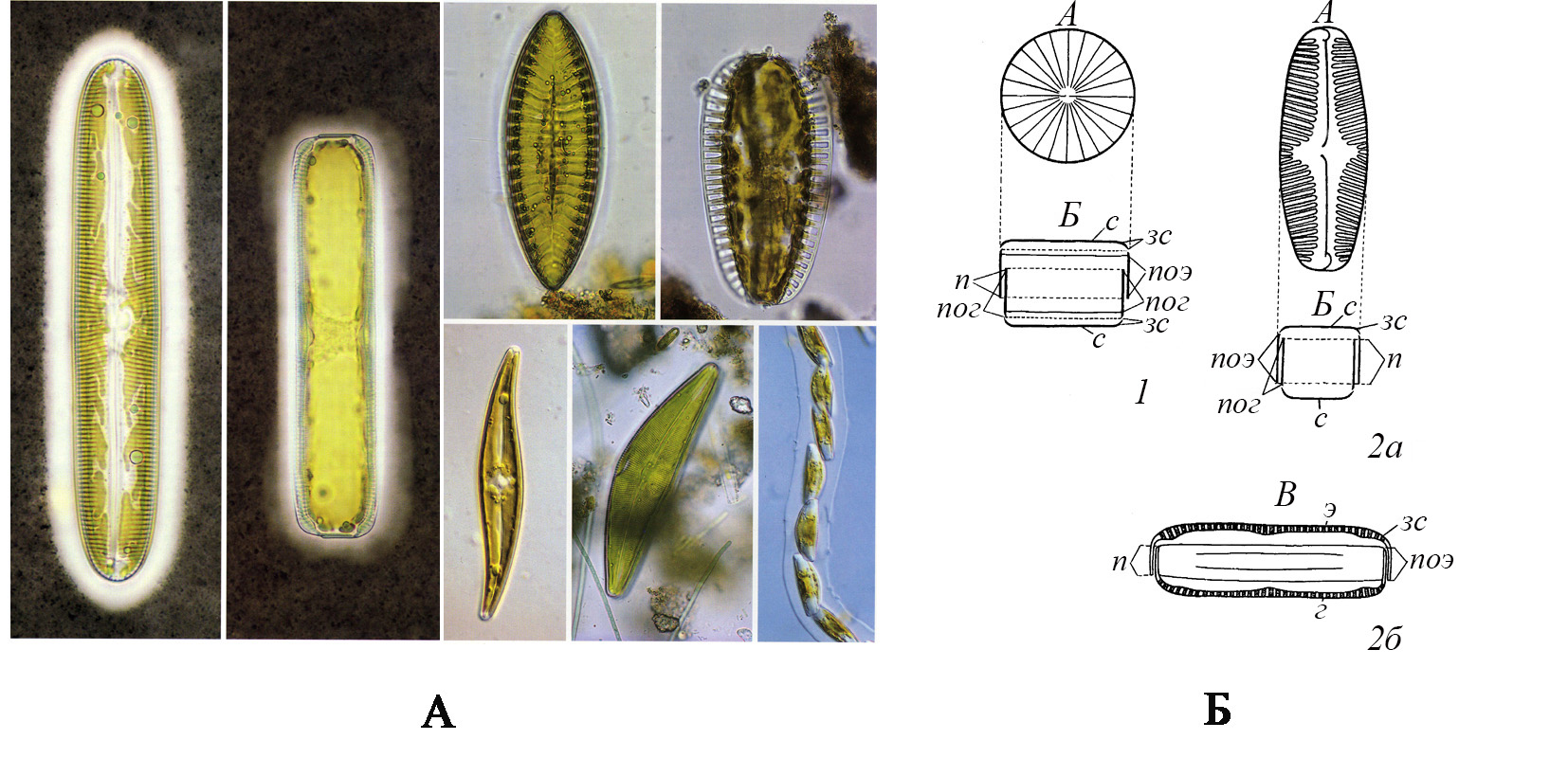
2. Биология, морфология, классификация 2.1. Диатомовые водоросли — эукариотические одноклеточные микроорганизмы с наружным панцирем, живущие одиночно или соединенные в колонии. Средние размеры их колеблются от нескольких микрон (мкм) до 50–100 мкм (гиганты достигают 2000 мкм, а самые малые имеют размеры около 1 мкм). Основная масса видов – фотосинтетики. Они имеют коричневато-желтую окраску, поскольку кроме хлорофилла а и хлорофилла с в хлоропластах содержится много каротиноидов (пигментов желто-коричневой окраски: фукоксантин, неофукоксантин, диадиноксантин, диатоксантин). Однако при недостатке света и в темноте некоторые виды могут переходить на гетеротрофный тип питания при условии достаточного количества органического углерода. Менее 10 видов облигатные гетеротрофы. Запасные вещества — масла, волютин, хризоламинарин, которые составляют до 20% сухого веса. Отличительная особенность этой группы – кремнеземный панцирь (гидрат окиси кремния SiO2.×nH2O), состоящий из двух створок (эпитеки и гипотеки) и пояска между ними (рис. 1). Толщина стенок панциря колеблется от 0,08 до 3 мкм. По весу панцирь составляет до 30–50% от сухого веса клетки. Кроме кремнезема в состав панциря входит примесь органических соединений и некоторых металлов (железо, алюминий, магний). Снаружи и изнутри панцирь покрыт тонким органическим слоем, состоящим из пектиновых веществ. Панцирь тонко перфорированный, перфорации занимают до 10–75% от площади поверхности клетки. Процесс образования панциря сложный и еще мало изученный, однако известно, что под плазмалеммой образуется трехслойная мембрана силикалемма, в которой возникают так называемые пузырьки – отложения кремнезема (silicon deposition vesicle – sdv). Силикалемма — активная избирательная мембрана, проходя через которую кремнезем концентрируется в пузырьке (sdv), его концентрация превышает концентрацию кремнезема в окружающей среде в 250 и более раз. Кремний, не считая кислорода и водорода, является в клетках диатомовых доминирующим элементом, превосходя по содержанию углерод. Основное количество кремнезема сосредоточено в панцире диатомовых, однако соединения кремния играют огромную роль не только как скелетный элемент, но и как элемент, необходимый для осуществления всех функций живого организма: питания, дыхания, роста и размножения. Кремний входит в состав не только цитоплазмы, но и органелл, особенно митохондрий. Именно панцири диатомовых хорошо сохраняются в ископаемом состоянии.
Рис. 1. Клетка и панцирь диатомей А – Общий вид клеток диатомей (по Carter-Lund, Lund, 1995).
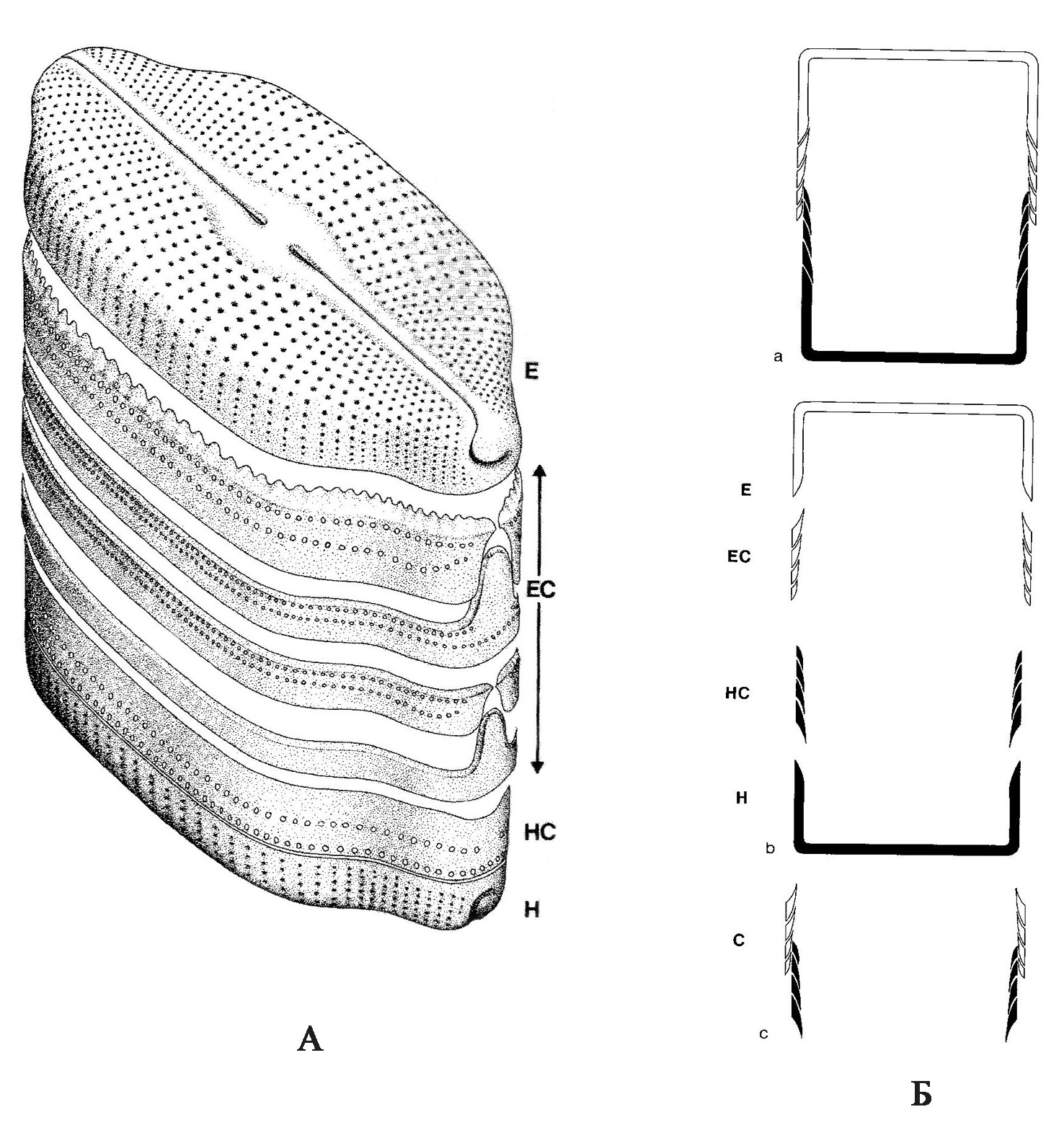
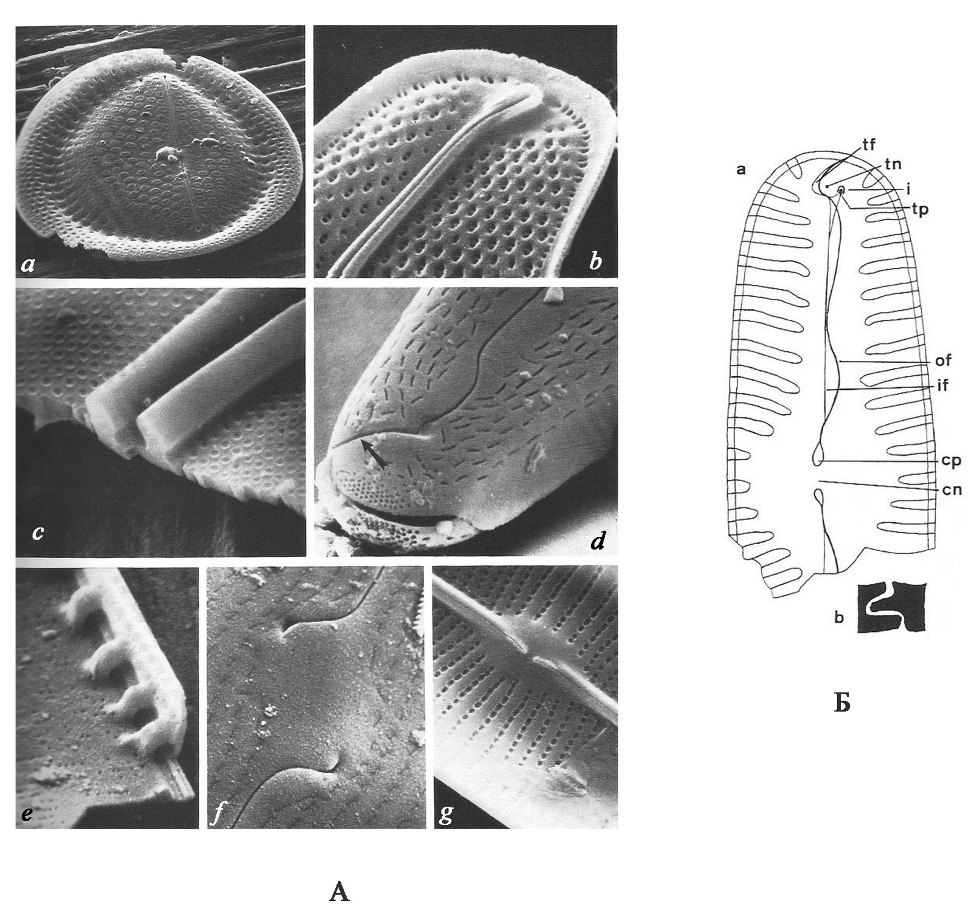
Воспроизведение[1]. Размножаются диатомеи вегетативным и половым путем. При вегетативном размножении клетки делятся пополам, и каждая дочерняя клетка получает одну бόльшую материнскую половинку панциря (эпитеку), а вторую меньшую (гипотеку) достраивает самостоятельно. В результате деления происходит мельчание клеток. Половой процесс протекает в виде изогамии, физиологической анизогамии и оогамии. Изогамия и анизогамия характерны для пеннатных диатомей. Оогамный половой процесс характерен для центрических и одного рода пеннатных диатомей. Сперматозоиды одножгутиковые, жгутиковый аппарат сильно редуцирован, женские гаметы – яйцеклетки, неподвижные. Жизненный цикл диплобионтный с гаметической редукцией. При образовании гамет происходит редукционное деление ядра, гаметы гаплоидные. В результате слияния гамет образуется зигота, которая затем начинает расти (т.е. становится ауксоспорой – спорой роста). Диатомовые водоросли – единственная группа среди растительных организмов, в жизненном цикле которых имеется стадия ауксоспорообразования. В зрелых ауксоспорах после слияния ядер (кариогамии) возникает диплоидное ядро. Ауксоспоры увеличиваются в объеме, достигая максимальных для вида размеров; затем внутри ауксоспоры образуется инициальная клетка: она приступает к вегетативному делению и дает начало новому клону (Давидович, 2017). 2.2. Классификация диатомовых основана на форме и строении панциря. Геометрическая форма панциря разнообразна: шар, диск, цилиндр, палочка, ладьевидная, S-образная. Разделение диатомовых на две большие группы основано на типе симметрии. Виды со створками с осью симметрии, через которую можно провести три и более плоскостей симметрии (актиноморфные) объединяют в центрические диатомовые (Centrophyceae); со створками, через которые можно провести не более двух плоскостей симметрии (зигоморфные) – в пеннатные диатомовые (Pennatophyceae). Настоящую «революцию» в диатомологии произвело широкое внедрение в практику исследований в 70-е годы XX века сканирующего и трансмиссионного электронных микроскопов. Их использование позволило получить новые и важные сведения по морфологии и строению панциря диатомей, что привело к изменению в значительной степени прежних представлений о его тонкой структуре. Были установлены особенности строения многих деталей панциря, недостаточно хорошо различимых в световом микроскопе, а также выявлены ранее неизвестные элементы структуры. Качественно новый уровень исследований с помощью электронного микроскопа значительно увеличил число признаков, которые можно использовать для систематики. Полученные данные привели к коренному пересмотру основных представлений о морфологии диатомей и их классификации. В настоящее время без изучения панцирей диатомей с помощью сканирующего электронного микроскопа невозможно описание, а часто и определение видов. В последние годы также привлекаются данные молекулярно-генетических исследований для установления родства и разграничения так называемых криптических (скрытых, не отличимых морфологически) видов. 2.3. Строение панциря диатомей и его структурные элементы[2] Панцирь состоит из двух половинок: верхняя бóльшая – эпитека и нижняя меньшая – гипотека (рис. 2). Каждая половинка состоит из эпивальвы (верхняя створка) и эпицингулюма (поясковый ободок эпитеки) и гиповальвы (нижняя створка) и гипоцингулюма (поясковый ободок гипотеки). Поясковые ободки, накладываясь друг на друга, образуют поясок (цингулюм). У створки различают лицевую поверхность створки (valve face) и загиб створки (valve mantle). Загиб – это краевая часть створки, отличающаяся наклоном, иногда также структурой. У диатомовых различают вид со створки и вид с пояска. Положение, при котором мы видим только створку, называется вид со створки; положение, когда видны соединение створок и поясок – вид с пояска или вид с загиба (в тех случаях, когда поясок отсутствует). У некоторых видов створки соединяются краями загибов, у них поясок появляется только во время деления клетки. У многих видов возникает вставочный ободок, который образуется между загибом створки и поясковым ободком. Вставочных ободков может быть много, но самый молодой образуется у загиба створки, а старый находится около пояскового ободка. Форма вставочных ободков может быть кольцевидной, воротничковидной и др. Функция этого элемента панциря заключается в увеличении объема и обеспечении роста панциря. От внутренних стенок вставочных ободков внутрь панциря у некоторых диатомей врастают тонкие неполные кремнеземные перегородки – септы, которые перегораживают клетку и увеличивают внутреннюю поверхность клетки диатомей. Через многочисленные перфорации панциря осуществляется связь с внешней средой. Перфорации имеют сложное строение и разнообразную форму, их называют порами, пороидами, ареолами и альвеолами в зависимости от формы и строения (рис. 3). Как правило, перфорации с одной стороны затянуты тонкой кремнеземной пластинкой, называемой велумом, который, в свою очередь, пронизан тончайшими крибральными порами, размеры которых менее 1 мкм.
Рис. 2. Схема строения панциря пеннатной (навикулоидной) диатомеи (по Round et al., 1990) А – общий вид: Е – эпивальва, ЕС – эпицингулюм, состоящий из четырех вставочных ободков, НС – гипоцингулюм, Н – гиповальва.
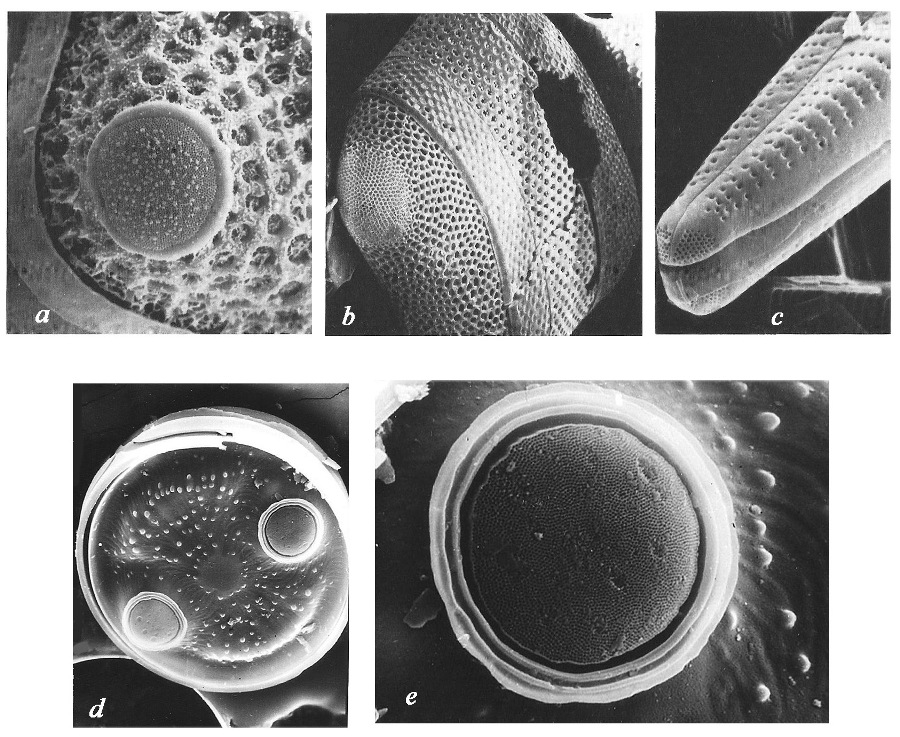
Среди основных структурных элементов створок диатомей выделяются следующие: Пора – сквозное, открытое с обоих концов отверстие в тонкой кремнеземной стенке панциря (створки). Поровый канал – узкий проход в толстостенном базальном слое кремнезема, обычно открытый на наружной и внутренней поверхности панциря. Ареола – регулярно повторяющаяся перфорация базального кремнеземного слоя, закрытая велумом на наружной и/или внутренней поверхности. Пороидная ареола или пороид – проход, который не расширяется в толще створки, имеющий цилиндрическую или слабо коническую форму, закрытый велумом на наружной, реже на внутренней поверхности створки. Локулярная ареола или локула – проход, который расширяется в толще створки, закрытый на наружной или внутренней поверхности створки. Диаметр форамена локулярной ареолы всегда меньше диаметра камеры, последняя расширена в толще базального слоя. Форма камер различна. Альвеола – удлиненная камера, выходящая из осевой или центральной части створки к краю, открывающаяся на внутренней поверхности створки большим отверстием, а на наружной поверхности перфорированным слоем. Велум – тонкий перфорированный слой кремнезема, закрывающий ареолу на наружной или внутренней поверхности панциря. Форамен – открытое отверстие, противоположное велуму. Гимен – тонкая кремнеземная мембрана, закрывающая у пеннатных диатомей ареолы.
Рис. 3. Перфорации стенки панциря диатомей А – Cхема строения перфораций: а – поры в базальном слое кремнезема, б – поровые каналы, в – пороидные ареолы, г – пороидная ареола с волой, д – локулярная ареола, бск – базальный слой кремнезема, в – вола, к – крибрум, кла – камера локулярной ареолы, п – пора, пк – поровый канал, ппа – проход пороидной ареолы, ф – форамен (по Николаев, Харвуд, 2002).
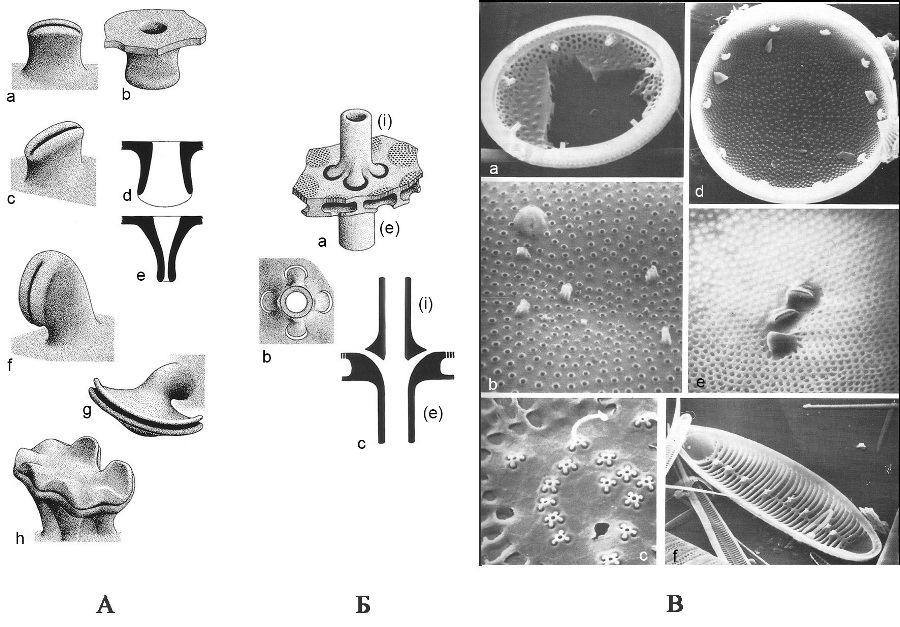
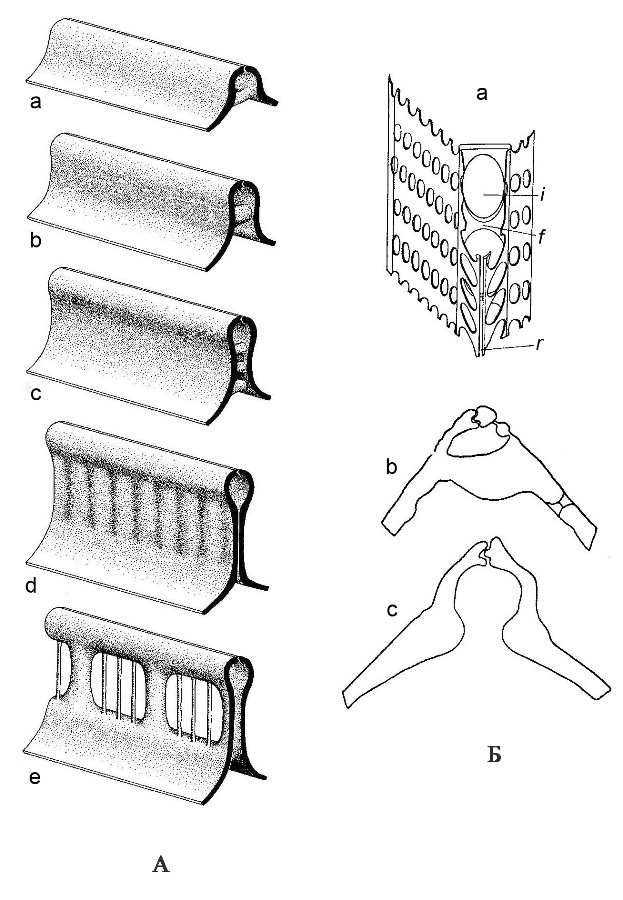
Выросты (portules). На створке имеются разнообразные перфорации в виде выростов, соединяющие протопласт с внешней средой и участвующие в экскреции слизи, состоящей из полисахаридов (рис. 4). У центрических и некоторых групп пеннатных диатомей (бесшовных и эуноциевых) имеются специализированные двугубые выросты – римопортулы (labiate process – rimoportula). Это структуры, которые на внутренней поверхности панциря имеют вид щели, окруженной гиалиновым утолщением, такая структура напоминает губы, откуда и произошло название. Створку пронизывает канал, который на наружной поверхности заканчивается трубкой или простым отверстием. На внутренней поверхности такая структура открывается воронкой, грибовидной структурой или подковообразной щелью. Различают сидячие двугубые выросты и на ножках (рис. 5). Другой тип трубчатых структур – специализированные выросты с опорами – фультопортулы (strutted process – fultoportula), известен только у одной группы центрических диатомей из талассиозировых. Фультопортула – это трубка, окруженная 2–5 камерами, или сопутствующими порами, и изолированными изогнутыми опорами. Считают, что эта структура (фультопортула) выделяет хитиновые фибриллы, которые способствуют формированию колоний, увеличивая плавучесть клеток, и защищают клетки от выедания.
Рис. 4. Типы выростов на створках диатомей (по Round et al., 1990) А – схема строения двугубых выростов: a–e – простой двугубый вырост (римопортула); a, c – вид с внутренней поверхности створки (видно «губовидное» отверстие выроста), b – вид с наружной поверхности створки (видно округлое поровидное отверстие), продольное (d) и поперечное (e) сечения выростов, показанных на фигурах a–c; f–h – примеры различной формы двугубых выростов с внутренней поверхности.
Рис. 5. Двугубые выросты и выросты с опорами a–d – двугубые выросты: a, b – щелевидный сидячий (род Hemiaulus); c – канал, который на наружной поверхности заканчивается трубкой (d) или простым отверстием (c). На внутренней поверхности такая структура открывается воронкой (c) (род Hemidiscus), или подковообразной щелью (род Aulacodiscus) (по Николаев, Харвуд, 2002); е – выросты с опорами: по краю – с двумя опорами, в центре – с тремя опорами (род Cyclotella) (ориг. Г.К. Хурсевич).
У некоторых колониальных или прикрепленных форм диатомей имеются особые структуры «глазки» (ocellus) и «ложные глазки» (pseudoocellus) и поровые поля у пеннатных диатомей. Глазок – это кремнеземная пластинка, пронизанная тесно расположенными мелкими порами – пореллами и окруженная утолщенным гиалиновым краем. Ложный глазок – участок створки с более мелкими ареолами по сравнению с ареолами на остальной части створки (рис. 6).
Рис. 6. Примеры особых структур створки в виде глазков и ложных глазков a – глазок (род Amphitetras), b – ложный глазок (род Isthmia), c – поровое поле (род Gomphonema) (по Round et al., 1990); d – пример створки (род Auliscus) с глазками и деталь (е) – глазок c пореллами (ориг. Т.Ф. Козыренко).
У пеннатных диатомей имеется особая структура – шов. Шов – удлиненная щель или две щели в стенке створки. Функция шва – обеспечение скользящего движения по субстрату. Считают, что через шов освобождается слизь, состоящая из фибриллярных полисахаридов, которые гидратируются, превращаются в тяжи и выделяются через шов на субстрат, обеспечивая движение клетки. Когда имеются две щели, они разделены центральным узелком. Расширения щели у центрального узелка и у апикального конца называются соответственно центральной и конечной порой. Различают щелевидный шов и каналовидный (рис. 7).
Рис. 7. Щелевидный и каналовидный швы на створке диатомей А – Система шва пеннатных диатомей: а – простая щель (род Anorthoneis); b – гребни вдоль щели шва на внутренней поверхности створки и губообразно утолщенное окончание внутренней щели (хоботок – helictoglossa) (род Pleurosigma); c – массивные гиалиновые утолщения вдоль шва (род Frustulia);
Щелевидный шов состоит из пары продольных щелей (ветвей шва), соединенных на середине створки центральным узелком и заканчивающихся близ концов створки конечным узелком. В стенке створки щель может быть коленчато-изогнутой. Щель (slit), открывающаяся наружу, называется наружной щелью, а открывающаяся в полость клетки – внутренней щелью. В центре створки выделяют центральную щель (central fissure) – это продолжение внутренней или наружной ветви щели, не пронизывающие створку насквозь и проходящие поверх центрального узелка. Конечная щель (terminal fissure) – это несквозное продолжение наружной щели шва, оканчивающейся в виде полулунной или слегка извитой терминальной щели. На концах створок на внутренней поверхности внутренняя щель формирует губообразное утолщение – хоботок (helictoglossa). Каналовидный шов имеет вид трубки, расположенной в толще створки, в гребневидном утолщении – киле или на крыловидном утолщении – крыле. С внешней средой он сообщается щелью, а в полость клетки открывается рядом отверстий (интерфибулы), отделенных между собою перегородками (фибулами) (Anonymous ..., 1975) (рис. 8).
Рис. 8. Схема строения каналовидного шва А – разные типы киля: a – простой мелкий киль (большинство в роде Nitzschia), b – более глубокий киль с фибулой на одном уровне (род Psammodictyon), c – глубокий киль с фибулами на нескольких уровнях (Entomoneis alata), d – сближенные две стороны киля, образующие узкие трубки, связывающие канал шва с полостью клетки (некоторые представители рода Entomoneis), e – образование промежутков между связывающими трубками и упрощение стенок киля (рода Surirella, Stenopterobia, Campilodiscus) (по Round et al., 1990);
Тип и строение структурных элементов, их количество (обычно измеряемое на отрезок, равный 10 мкм) и характер расположения на створке имеют важное значение при классификации диатомей. Например, при описании центрических диатомей большое внимание уделяется определению диаметра створки, характеру расположения ареол на поверхности створки (ареолы могут быть сомкнуты в сеть или быть изолированы бесструктурными гиалиновыми участками, располагаться в радиальных, линейных, тангентальных и концентрических рядах, группироваться в пучки и т.д.), а также количеству ареол в 10 мкм. 2.4. Жизненные формы Диатомовые в своей основе одноклеточные организмы, но многие формы образуют колонии, в которых клетки соединяются с помощью различных приспособлений. Можно выделить следующие типы соединения клеток: с помощью соединительных шипов (linking spines) (например, у родов Aulacoseira, Cymatoseira), соединительных кремнеземных структур (siliceous structures) – например, у рода Rhizosolenia, щетинок (у рода Chaetoceros) и выростов-рогов (у рода Hemiaulus). Иной способ соединения – с помощью слизистых подушечек (mucilage pads) (у родов Grammatophora, Synedra, Amphitetras), или стебельков (stalks) (у рода Licmophora) (рис. 9). По образу жизни выделяют планктонные формы, пассивно парящие в толще воды, и бентосные, жизнь которых связана с субстратом. И планктонные, и бентосные формы могут быть одиночными или колониальными. Одиночные бентосные формы способны ползать по субстрату. Движение диатомей процесс сложный, существует несколько гипотез о механизме движения. Это может быть движение с участием слизистых веществ, освобождающихся через шов, или за счет трения о субстрат цитоплазмы, циркулирующей в шве и перемещающей клетку в направлении, противоположном току цитоплазмы.
Рис. 9. Некоторые типы соединения клеток диатомей в колонии (по Кроуфорд, Гибшубер, 2006) А, Б – с помощью специальных соединительных шипов (А – род Aulacoseira, Б – род Cymatoseira),
3. Методика обработки и изучения материала Изучение современных и ископаемых диатомей включает несколько этапов. Первый из них – сбор материала в природе и хранение материала в лаборатории. От точности записи (где, когда и как) собран материал, зависит дальнейшая интерпретация результатов. При сборе современного материала его необходимо фиксировать (Диатомовый анализ, 1949). Для этого обычно используют 40% формалин. При сборе каменного материала (осадочных пород) необходимо соблюдать максимальную осторожность и аккуратность, чтобы избежать заноса — поскольку размеры диатомей очень малы (микроны), их остатки легко могут быть занесены из одной пробы в другую. В целом, лабораторная обработка слагается из следующих основных этапов: 1) дезинтеграция породы путем проваривания в 30% растворе перекиси водорода (H2O2), или пирофосфате натрия (Na4P2O7×10H2O); 2) обогащение глинистых пород с тяжелой жидкостью удельным весом 2,6 – йодистой или кадмиевой (ICd2KI); 3) отмучивание и разделение осадка на фракции центрифугированием (при необходимости); 4) приготовление постоянных препаратов с использованием особых смол с показателем преломления выше n = 1,43: чаще всего употребляются анилин-формальдегидная смола («среда Эльяшева»), показатель преломления n = 1,68 и «Нафракс» (Naphrax) ― с показателем преломления n = 1,74; 5) просмотр под микроскопом и определение видов, анализ таксономического состава, сравнение с литературными данными и написание заключения. При определении видов важными показателями являются размерные характеристики створок, для чего пользуются окуляр-микрометр (цену деления окуляр-микрометра вычисляют с помощью объекта-микрометра при разных увеличениях). Измеряют линейные размеры объекта и количество структурных элементов (ареол, выростов и т.д.) в 10 микронах (Диатомовые водоросли СССР ..., 1974).
4. Экология и биогеография Диатомовые водоросли экологически очень разнообразная группа. Они распространены в планктоне и бентосе морских и континентальных водоемов, в почве (геофитон), на влажных субстратах, приподнятых над поверхностью земли (аэрофитон), во льдах (криофитон). Результаты многолетних наблюдений диатомовых водорослей позволили создать экологическую классификацию диатомей по их отношению к разнообразным факторам внешней среды. Выделяют систему галобов, в основе которой лежит отношение диатомей к солевому составу и солености, систему ацидобионтности – отношение диатомей к кислотности среды, система сапробионтности – отношение диатомей к загрязнению органическими веществами. Составлены списки характерных видов для той или иной экологической группировки. Это позволяет использовать систему индикаторных видов для определения экологической обстановки в ныне существующих местах обитания и для воссоздания палеообстановок прошлых геологических эпох (Диатомовый сборник, 1953; Van Dam et al., 1994; Баринова и др., 2000; и др.). В современных водоемах диатомовые, как правило, являются доминирующей группой, от которой зависит продуктивность водоема. В Мировом океане особенно велика роль планктонных диатомей, среди которых обычно различают две большие группы: 1) голопланктон, представители которого характерны для открытого океана (моря) и проводят в нем все стадии жизненного цикла; 2) неритический планктон - формы, характерные для относительно мелководных морских зон, на дне которых они в ряде случаев могут проводить часть своего жизненного цикла (Диатомовые водоросли СССР …, 1974). В морских экосистемах планктонные диатомеи составляют по биомассе около 90% массы всех водорослей и являются важнейшим звеном в цепочке пищевых связей, так как представляют основной источник питания для зоопланктона, создавая его пищевой ресурс. Таким образом, часть солнечной радиации, ассимилированная в процессе фотосинтеза, переходит в энергию органических соединений и затем передается с одного трофического уровня на другой. В целом, диатомовые продуцируют до 50% общего органического вещества планеты (Николаев и др., 2001).
Рис. 10. Продукция биогенного кремнезема в океанах в г/м2/год (абсолютные массы за год, определенные по первичной продукции Cорг. и отношению SiO2 аморфн./Cорг. во взвеси)
В современных бассейнах происходит непрерывный круговорот кремния, в котором диатомеи являются основными продуцентами биогенного кремнезема. В континентальных водоемах для построения панциря они извлекают кремнезем, который в основном привносится из почв, а также освобождается при выветривании горных пород. В океанах и морях наиболее распространенными кремнистыми организмами во взвеси морских вод являются планктонные диатомовые водоросли, а пояса кремнистой взвеси совпадают с поясами массового развития диатомовых (рис. 10, 11). Развитие и степень продуктивности диатомовых, а также значения биомассы диатомей в морских бассейнах неоднородны, так как они находятся в зависимости от насыщенности поверхностных вод биогенными веществами (кремниевой кислотой, а также нитратами, фосфатами и железом), необходимыми для жизнедеятельности клеток. Распределение этих элементов неравномерно в разных районах Мирового океана. Наиболее массовое развитие диатомей происходит в тех зонах, поверхностные воды которых характеризуются высоким содержанием упомянутых питательных веществ. Исходя из данных по значениям концентрации аморфного кремнезема, а также первичной продукции, в океанах выделяются три основных пояса высокой продуктивности: два в высоких-средних широтах и один (более слабый) в экваториальной области (рис. 11). Так как в целом, концентрация свободного кремнезема в поверхностных водах ниже, чем в глубинных, из-за того, что он вместе с другими минеральными солями постоянно извлекается планктоном, для продуктивного развития микроорганизмов необходимы новые поставки биогенных веществ. Наибольшее обогащение фотической зоны этими элементами осуществляется за счет подъема к поверхности глубинных вод, в которых имеется огромный запас минеральных солей. В современных океанах из-за термической стратификации водной толщи с наличием термоклинного барьера, такой подъем возможен только в районах интенсивного вертикального перемешивания вод. К таким районам, в первую очередь, относятся зоны, совпадающие с центральными частями больших океанических круговоротов, где происходит перемешивание вод за счет дивергенций и циклонических круговоротов. Подъем глубинных вод осуществляется также в районах региональных апвеллингов, приуроченных к прибрежным зонам западных окраин материков, в результате сгонных явлений, в конечном итоге, за счет силы Кориолиса (Kennett, 1982; Barron, 1993). Высокая продуктивность кремнистого планктона характерна также для апвеллинговых зон, находящихся на стыке разнонаправленных поверхностных течений в экваториальных районах восточных частей океанов. Самые же высокие концентрации свободного кремнезема и минеральных солей, а соответственно и наибольшая продуктивность диатомовых, характерны для холодноводных областей океанов приантарктических районов, а также Северной Пацифики и Северной Атлантики (рис. 11). В целом, в этих областях разница между температурами поверхностных и глубинных вод не очень значительна. Поэтому отсутствие резкого термоклина делает возможным интенсивное вертикальное перемешивание.
Рис. 11. Скорость экстракции растворенного кремнезема (в г/м2/год) фитопланктоном в поверхностных водах разных районов Мирового океана (по Lisitzin, 1972; Calvert, 1974)
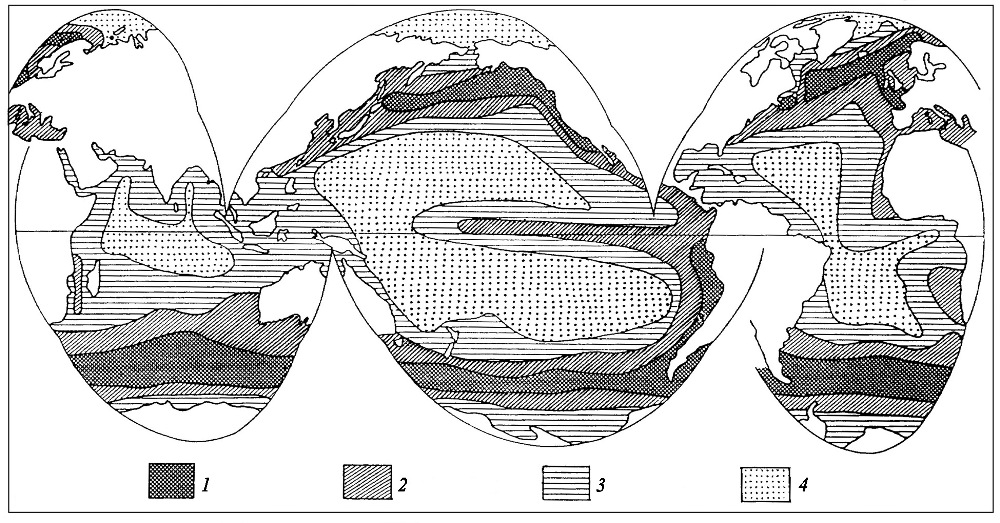
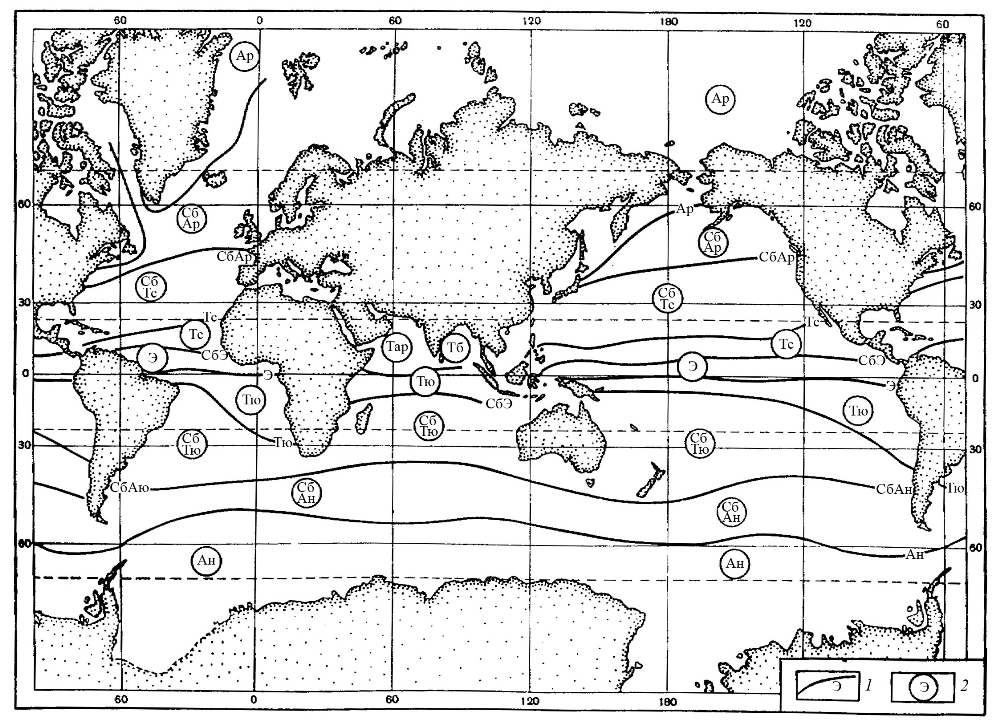
Таксономический состав диатомовой флоры в первую очередь зависит от температуры поверхностных вод, то есть от климатической зональности, когда различным географическим зонам соответствует флора определенного видового состава. В зависимости от положения климатических зон и деятельности течений в океанах существуют различные поверхностные водные массы, которые характеризуются определенными режимами температур и солености. В целом, в современном Мировом океане различаются следующие поверхностные водные массы: экваториальные, тропические, умеренные (субтропические и субполярные) и полярные (арктические и антарктические (рис. 12). Они разделены переходными – фронтальными зонами (фронтами), которые простираются субширотно в соответствии с границами климатических зон. Во фронтальных зонах развиты диатомовые сообщества смешанного состава, то есть переходные между комплексами соседних водных масс. Фронты служат термическими барьерами для миграции планктонных диатомей, поэтому распространение многих видов ограничено определенной поверхностной водной массой, то есть являются наиболее для нее показательными. Для каждой поверхностной водной массы типичны свои показатели температуры, солености, плотности, скорости горизонтального и вертикального перемешивания, содержания биогенных элементов, кислорода и проч. Этим массам соответствуют ассоциации диатомей определенного видового состава с разными доминантами. Кроме того, распространение некоторых видов ограничено определенной поверхностной водной массой, то есть являются наиболее для нее показательными (индикаторными). В течение года в пределах одной водной массы обычно происходит также смена доминирующих в ассоциациях видов, что вызывается сезонными изменениями температуры вод, освещенности и степени насыщенности минеральными веществами. В частности, сезонные апвеллинги могут быть причиной интенсивного развития определенных форм.
Рис. 12. Поверхностные океанические водные массы и фронты (по Степанов, 1974) 1 – океанические фронты: Э – экваториальный, СбЭ – субэкваториальный, Тс – северный тропический, Тю – южный тропический, СбАр – субарктический, СбАн – субантарктический, Ар – арктический,
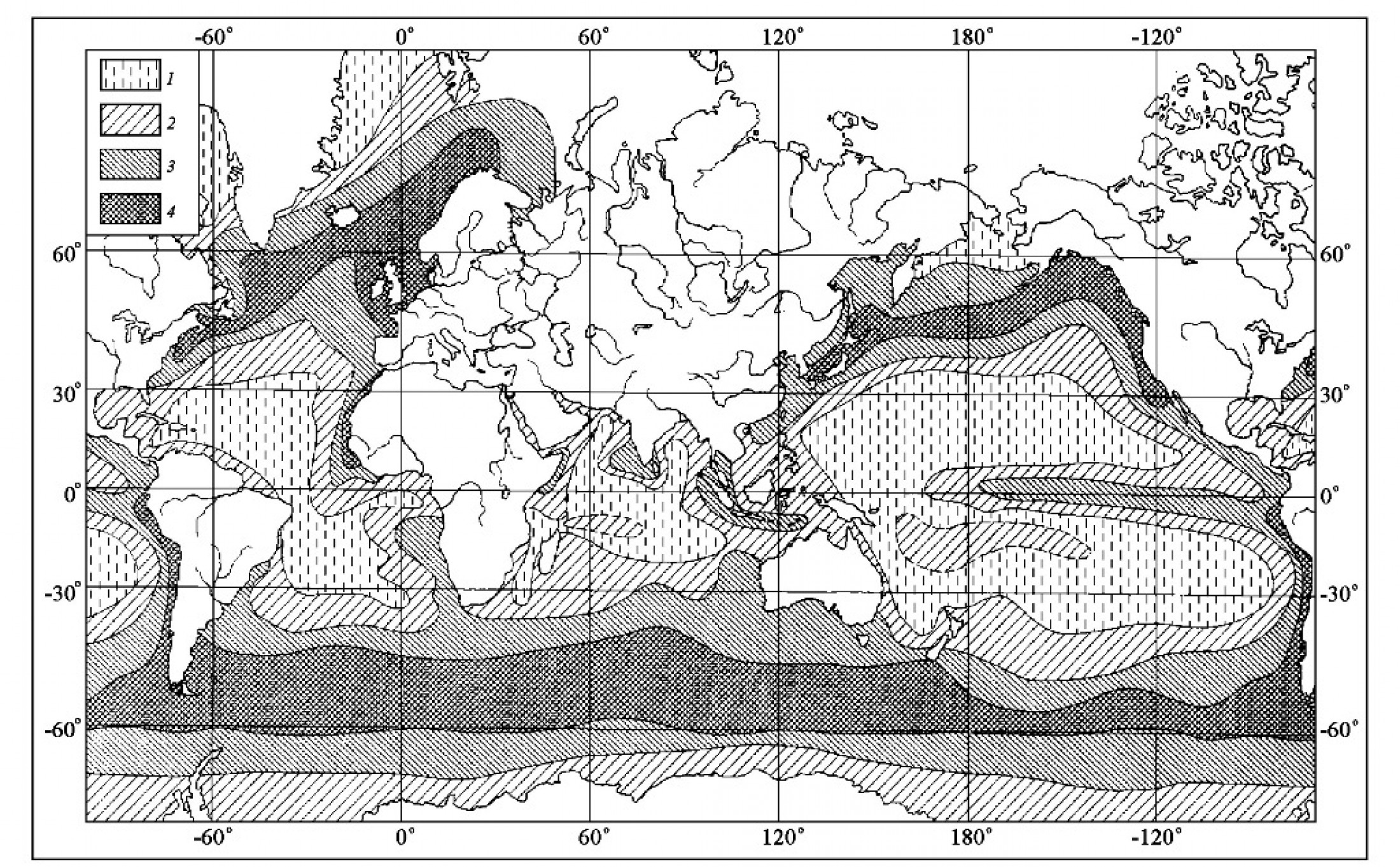
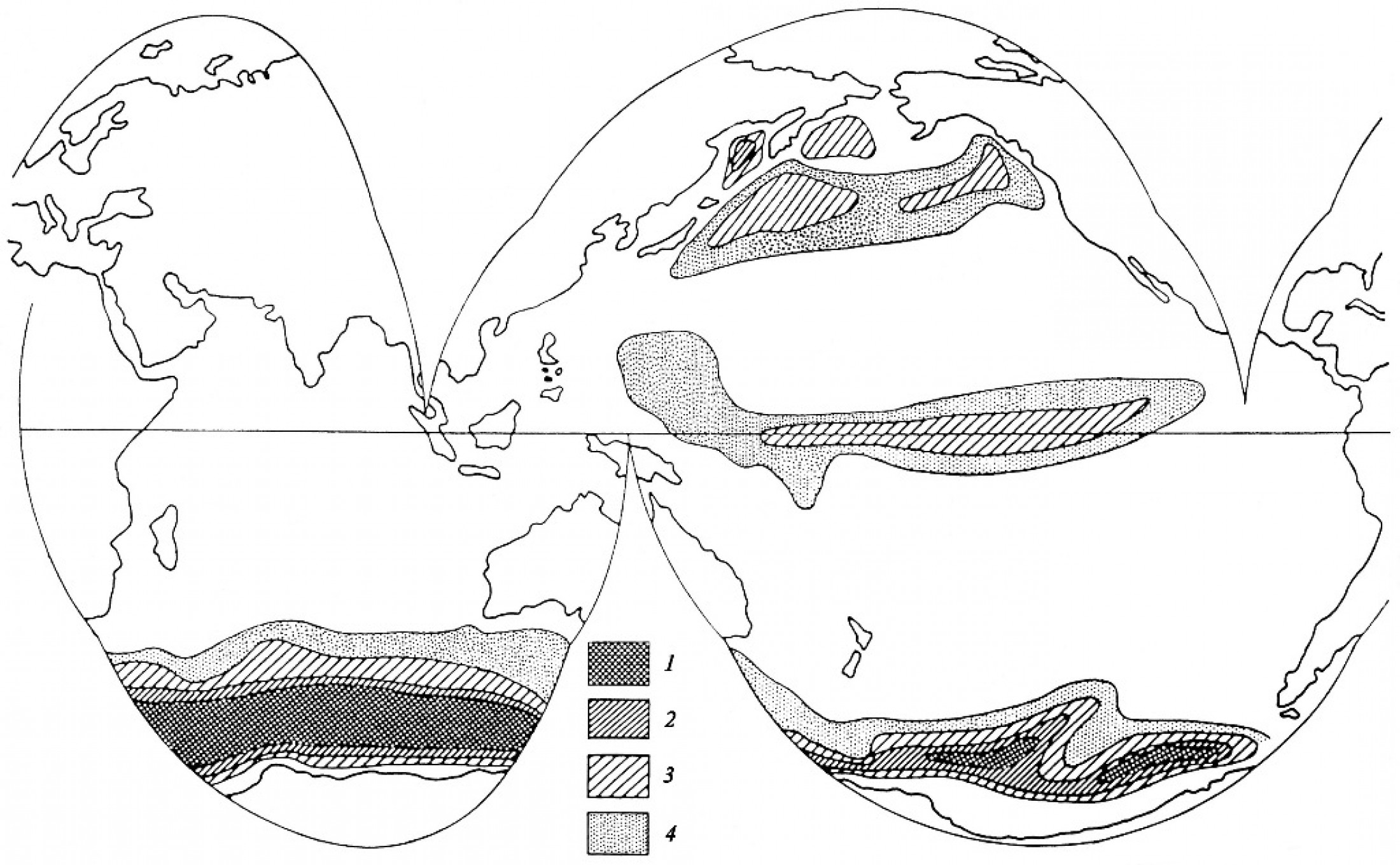
5. Роль диатомей в осадконакоплении и породообразовании Диатомовые водоросли являются не только основными продуцентами биогенного кремнезема, но и главными его поставщиками в донные осадки при седиментогенезе. В континентальных водоемах поставка кремнезема в осадки практически полностью осуществляется за счет пресноводных диатомей. Что касается современного морского кремненакопления, то более 90% в нем приходится на долю диатомовых (Лисицын, 1978). В Мировом океане прослеживаются три основных пояса современного осадочного кремненакопления: 1) южный, охватывающий почти сплошной полосой южное полушарие между примерно 45–65º ю.ш.; 2) приэкваториальный и 3) северный, развитый в северной части Тихого океана (включая Берингово и Охотское моря) и Северной Атлантике (рис. 13). Наибольшая ширина пояса диатомовых илов и самые высокие содержания кремнезема в осадках (с образованием диатомовых илов) характерны для южного полушария где накапливается более 80% биогенного кремнезема Мирового океана. Положение поясов осадочного кремненакопления в целом совпадают с таковым регионов, поверхностные воды которых характеризуются наибольшей продуктивностью диатомей. Это отражает общую взаимосвязь между продуктивностью диатомовых и их участием в формировании кремнистых осадков.
Рис. 13. Распространение и процентная концентрация биогенного опала в поверхностных осадках Тихого и Индийского океанов (по Lisitzin, 1972; Calvert, 1974)
При формировании разных типов донных осадков с участием остатков диатомовых водорослей большое значение имеют условия и режим, при котором происходит процесс осадконакопления. В зависимости от типа седиментогенеза в различных фациальных областях океана диатомеи принимают участие в формировании разных типов осадков. В продуктивных зонах панцири диатомей поступают на дно практически непрерывно и с достаточно большой скоростью, накапливаясь в осадках в массовом количестве. В результате, в таких районах могут формироваться и накапливаться диатомовые илы, которые занимают на площади дна Мирового океана несколько десятков миллионов квадратных километров. Образование диатомовых илов происходит в тех районах, где количество попадающих на дно бассейна панцирей диатомей выше темпа поступления другого осадочного материала, как терригенного, так и органогенного. Ярким примером служат диатомовые илы, накапливающиеся в глубоководных районах открытого океана вблизи Антарктиды. В других обстановках седиментации, при развитии диатомей в морском бассейне, во многих случаях они в том или ином количестве захорониваются и в осадках другого генезиса – как терригенных, так и органогенных. Достоверные находки наиболее древних морских диатомовых известны из отложений нижнего мела, а пресноводных – из толщ палеоцена (Gersonde, Harwood, 1990; Николаев и др., 2001; Harwood et al., 2007; Siver et al., 2016). Породообразующее значение диатомеи приобретают, начиная с позднего мела. С этого времени получают распространение морские диатомиты – кремнистые осадочные породы, образованные в основном панцирями диатомей, а также туфодиатомиты, диатомовые глины и сланцы - многокомпонентные породы, в состав которых помимо органогенно-кремнистой составляющей входит пирокластический, терригенный и пелитовый материал. Эти породы по существу являются ископаемыми аналогами диатомовых и диатомово-глинистых илов и близких к ним морских осадков. Наиболее древние озерные диатомиты известны, начиная с олигоцена.
6. Палеоэкология и реконструкции условий окружающей среды прошлого Как было сказано, диатомовые водоросли являются хорошими индикаторами параметров окружающей среды, в частности, режима температуры, солености, степени обогащенности питательными веществами, глубины бассейна и т.д. Поэтому изучение ископаемых комплексов диатомей в осадочной летописи предоставляет важную информацию при проведении палеоэкологических, палеоклиматических и палеогеографических реконструкций, а также выявлении перестроек в экосистемах. В большинстве случаев при этом используется принцип актуализма. Вот почему для различного рода реконструкций необходимо учитывать данные по особенностям распределения современной диатомовой флоры в зависимости от климатической зональности и физико-химических характеристик вод, которые лежат в основе биогеографического, и в частности, фитогеографического, районирования. Выводы, вытекающие из тщательного изучения связей и изменений в современных экосистемах, представляют большую важность при палеоэкосистемном анализе. Например, анализ количественных соотношений тепловодных и холодноводных видов используется для оценки палеотемператур фотической зоны вод. В четвертичной геологии данные диатомового анализа широко используются при изучении ледниковых и межледниковых периодов. Анализ таксономического состава разновозрастных древних морских диатомовых ассоциаций позволяет делать выводы о распространении различных поверхностных водных масс и смещении их границ во времени, а также степени контраста термических градиентов как барьеров для миграций планктона. Наряду с палеоклиматическим аспектом, большую ценность имеют данные по палеогеографии, так как анализ ареалов дает возможность судить о масштабе и путях миграций планктонных видов. Характер распространения биогенных кремнистых осадков дает представление о степени продуктивности диатомовой флоры в том или ином регионе в соответствующий интервал геологического времени. В последние десятилетия широкое применение находит как мониторинг изменений таксономического состава диатомей в водах внутриконтинентальных водоемов, так и детальное изучение диатомовых в позднеголоценовых осадках этих бассейнов. Результаты исследований дают возможность делать выводы о степени влияния антропогенного воздействия на развитие водоема, в частности, позволяют оценивать степень его промышленного загрязнения. Однако при изучении материала в донных осадках и осадочных породах, а также использовании полученных данных для проведения реконструкций условий окружающей среды прошлого, часто имеются определенные ограничения и трудности. Прежде всего, они связаны со степенью сохранности панцирей древних диатомей.
7. Сохраняемость в донных осадках и осадочных породах Во многих случаях таксономический состав диатомей в биоценозах фотической зоны и в танатоценозах донных осадков этого же района не одинаков. Прежде всего, это касается глубоководных морских бассейнов. Одним из основных факторов, который определяет состав танатоценозов, является выборочная сохранность створок диатомей в осадках, обусловленная растворением. Так как в целом морские воды недонасыщены свободным кремнеземом, процесс растворения начинается практически сразу при опускании панцирей на дно сквозь толщу. Поэтому, чем длительнее промежуток времени воздействия морской воды на панцири при их осаждении, тем более вероятно их растворение. Створки диатомей имеют разную толщину и степень окремнения, большая их часть растворяется, не попадая в осадок. В результате виды с хрупкими и тонкими створками могут практически полностью отсутствовать в составе танатоценозов, тогда как формы с более толстостенными сильно окремненными створками имеют лучшие шансы уцелеть при осаждении. То есть во многих случаях отмечается несоответствие между количественным и видовым составом диатомей в планктоне и осадках, что часто приводит к тому, что массовые виды в них не одни и те же. Попадая на дно бассейна в осадки, при отсутствии длительного воздействия щелочной среды поровых вод со значениями pH более 7, панцири в целом больше не подвергаются активному растворению. Таким образом, полного соответствия между составами биоценозов и танатоценозов практически никогда не наблюдается. Однако, как показали результаты многих исследований, несмотря на выборочное растворение и другие процессы, распределение диатомей в поверхностном слое осадков во многих случаях близко отражает закономерности, которые свойственны их распределению в современном фитопланктоне. Последующему дополнительному воздействию панцири подвергаются при захоронении осадков и образовании из них горных пород. При диагенезе и катагенезе степень сохранности створок может ухудшаться вплоть до полной перекристаллизации, то есть изменению первично-органогенной природы кремнезема. Это, прежде всего, связано с действием процессов, проходящих в определенных термодинамических (PT) условиях на глубине.
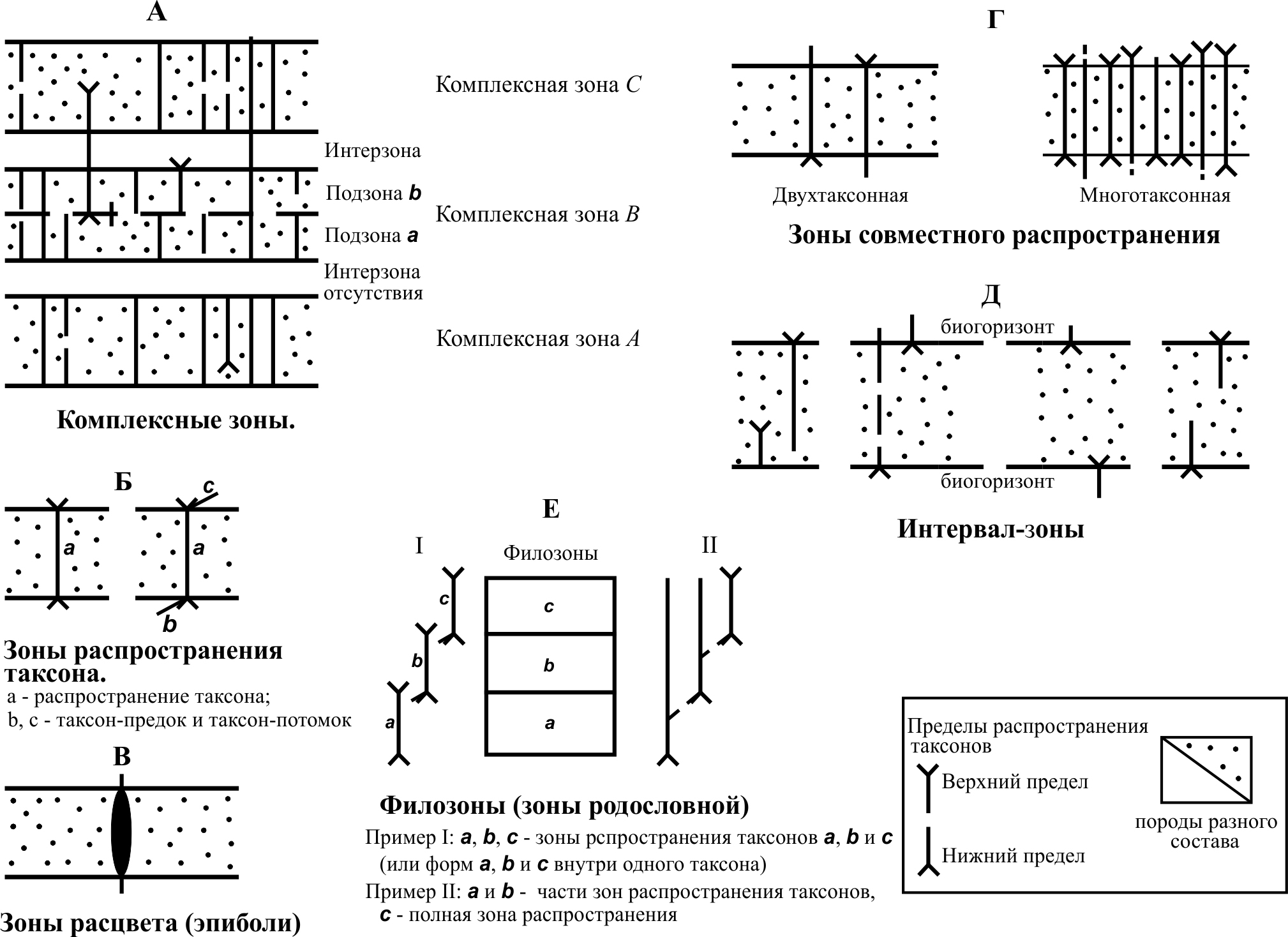
8. Значение для биостратиграфии 8.1. Диатомеи начали использовать в стратиграфии в первой половине XX века. Среди пионеров в этой области были российские ученые В.С. Порецкий, А.П. Жузе и В.С. Шешукова-Порецкая, а также американцы Дж.Д. Хэнна (G.D. Hanna) и К.Е. Ломэн (K.E. Lohman), которые одними из первых показали принципиальную возможность использования диатомовых при корреляции кайнозойских отложений удаленных разрезов. В результате изучения ряда наземных разрезов и образцов донных отложений в них были выделены палеонтологические ассоциации разного возраста, и предложен ряд биостратиграфических подразделений по диатомеям. В первую очередь, они представляли собой горизонты и слои с флорой. Значительный интерес к изучению диатомовых водорослей проявился в конце 60-х годов прошлого века, когда в Мировом океане начали широкомасштабно проводить глубоководное бурение. При изучении кернов скважин было выявлено, что во многих случаях в них в большом количестве представлены остатки диатомей, а также, что они с успехом могут использоваться в качестве одного из основных «инструментов» для проведения расчленения и корреляций вмещающих отложений на детальной (зональной) основе. Обработка материалов глубоководных скважин дала возможность проследить в относительно полных разрезах пелагических фаций различных регионов последовательную смену комплексов, оценить интервалы стратиграфического распространения многих видов диатомей (включая, впервые описанные), а также выявить стратиграфически важные таксоны. Это позволило подойти к разработке океанических зональных шкал по диатомеям. Зоны представляют собой более дробные стратиграфические подразделения по сравнению с ярусами. Обычно они выделяются на биостратиграфической основе с учетом этапности эволюции ископаемых организмов. Под зоной (по диатомеям) понимаются отложения, сформировавшиеся за время существования определенного палеонтологического комплекса, отражающего этап эволюционного развития группы ископаемых (диатомовой флоры) и отличающегося от комплексов смежных слоев. При выделении зон и проведении их границ используются датированные уровни – прежде всего, уровни появления или исчезновения отдельных маркирующих видов. Возраст и изохронность таких уровней контролируются, в первую очередь, их корреляцией в разрезах с магнитостратиграфической шкалой и подразделениями по карбонатному планктону. Критерии проведения границ могут быть неодинаковыми, поэтому, в зависимости от этого, выделяются различные типы зон: комплексные зоны, зоны распространения (включающие зоны совместного распространения, зоны распространения таксона, филозоны), интервал-зоны и акме-зоны (рис. 14). При этом различия в составах сменяющих друг друга зональных сообществ, в основном, проявляются в ограниченном числе видов. Как показала практика, наиболее эффективные результаты по выделению зон достигаются в результате анализа стратиграфического распространения отдельных видов (необязательно относящихся к одному роду) или реже – родов. Именно применение такого подхода позволяет в итоге получать дробные и смыкающиеся биостратиграфические подразделения с относительно изохронными границами. При работе с морскими отложениями наибольшие интерес и ценность представляют зоны, выделенные по планктону, которые обеспечивают высокую разрешающую способность. Так как ассоциации планктона быстро эволюционируют и имеют широкое географическое распространение, их изучение позволяет успешно осуществлять дробное расчленение и обоснованные широкие корреляции древних толщ.
8.2. Расчленение мела Меловые диатомеи являются наиболее слабо изученными. Достоверные находки наиболее древних диатомовых водорослей известны из морских отложений нижнего мела (апт-альб). Раннемеловая флора с наилучшей степенью сохранности изучена из разреза глубоководной скважины, пробуренной в море Уэделла (Западная Антарктида) и датирована поздним аптом – ранним альбом в пределах 115–110 млн лет (Gersonde, Harwood, 1990). Одна из первых сводок о меловых диатомеях с обобщением и описанием имевшегося материала по верхнему мелу была опубликована в 70-х годах XX века (Стрельникова, 1974). Большой массив новых данных по морфологии, таксономии, биостратиграфии и эволюции меловых диатомовых получен за последние 20–25 лет, прежде всего, в результате совместных исследований В.А. Николаева (Россия) и Д.М. Хэрвуда (США). Однако при выделении стратиграфических подразделений меловых толщ по диатомеям до сих пор приходится сталкиваться с рядом проблем. Одна из них заключается в том, что диатомовая флора удовлетворительной сохранности известна из очень ограниченного количества местонахождений и характеризует отдельные интервалы разрезов (в основном, наземных). Однако в начале 2000-х годов результаты изучения разновозрастных комплексов в разрезах Канадской Арктики с привлечением данных по Северному Зауралью позволили предложить зональную схему по диатомеям для расчленения верхнего мела, включающую пять зон по диатомеям (Tapia, Harwood, 2002) (рис. 15).
Рис. 15. Зональная шкала по диатомеям для расчленения верхнего мела (по Tapia, Harwood, 2002)
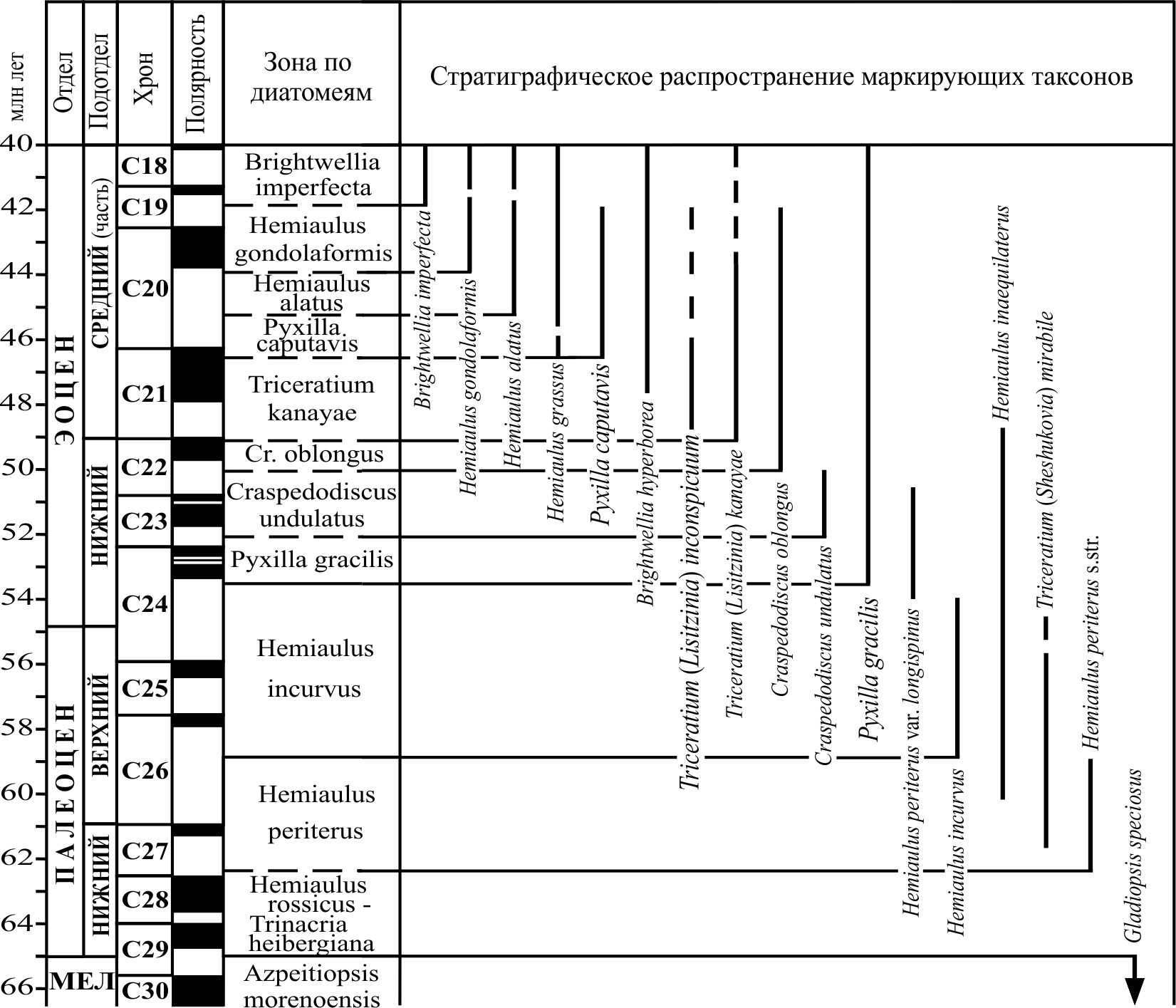
8.3. Зональные шкалы палеоцена – нижнего эоцена По сравнению с меловыми, диатомеи палеоценового и раннеэоценового возраста представлены в наземных разрезах и разрезах глубоководных скважин более широко. В частности, они развиты в отложениях древних эпиконтинентальных морей, где нередко имеют породообразующее значение. Однако основная сложность при изучении нижнего палеогена заключается в прослеживании последовательной смены комплексов в непрерывных разрезах. Как правило, диатомеями охарактеризованы только отдельные интервалы, и если говорить о нижнем палеогене в целом, то смыкаемость разработанных по диатомеям зональных подразделений для всего этого интервала отсутствует, а их сопоставления проводятся опосредованно. Шкалы палеоцена – нижнего эоцена разработаны с определенной долей условности, а предложенные для разных районов схемы расчленения находят пока, главным образом, локальное применение. На рис. 16 показаны подразделения океанической шкалы для палеоцена – нижнего эоцена, выделяемые в качестве зональных. Несмотря на несовершенство зон, разработанных по материалам океанических разрезов, они позволили значительно дополнить и уточнить существовавшие ранее схемы. Во-первых, была выявлена последовательность появления и исчезновения важных в стратиграфическом отношении форм морских диатомей в непрерывных разрезах отдельных интервалов палеоцена – нижнего эоцена разных районов океана. Соответственно на основе анализа датированных уровней были предложены биостратиграфические подразделения. Во-вторых, часть таких уровней напрямую скоррелирована с зонами по карбонатному планктону и динофлагеллатам, что позволило достаточно достоверно оценить возраст и объем выделенных зон или слоев с флорой. В-третьих, была выяснена большая степень сходства таксономического состава комплексов диатомей из разрезов нижнего палеогена океанических толщ и разрезов суши. Это сделало возможным использование в наземных разрезах (в которых прямые корреляции с ассоциациями карбонатного планктона и палеомагнитной шкалой в большинстве случаев отсутствуют) общих с океаном форм с установленными интервалами стратиграфического распространения.
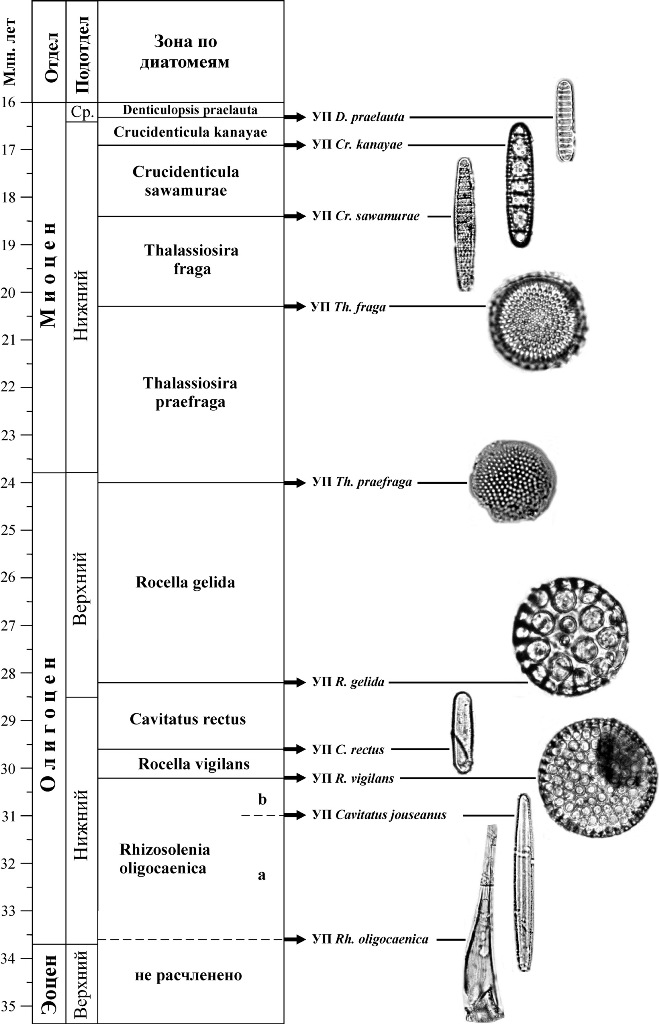
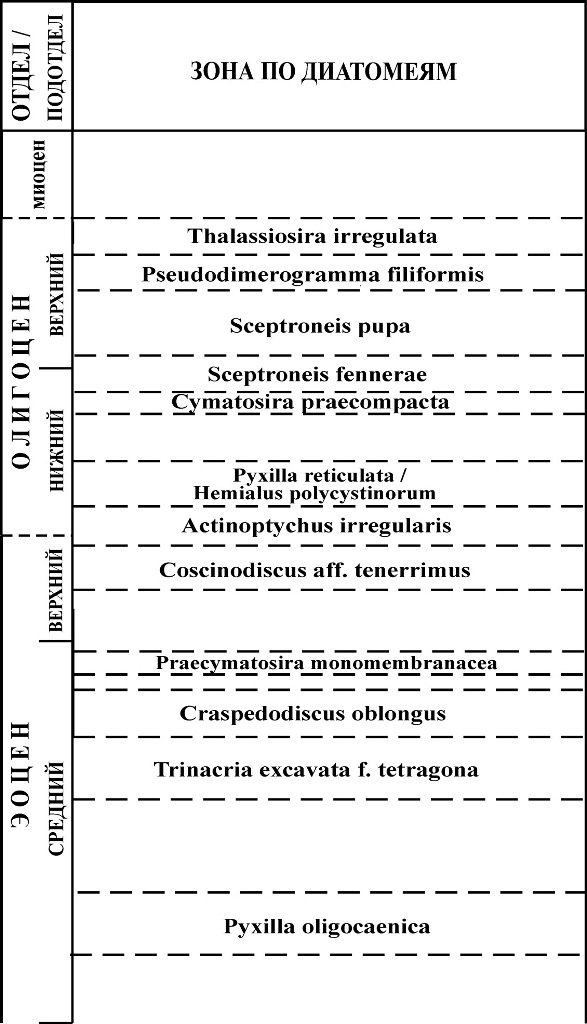
8.4. Зональные шкалы среднего эоцена-олигоцена Морские отложения среднего эоцена-олигоцена, охарактеризованные диатомовыми, имеют достаточно широкое распространение. Зональность для этого интервала разработана лучше по сравнению с более древними толщами. Результаты исследований диатомовых комплексов в разрезах скважин глубоководного бурения с привлечением материалов по наземным разрезам привели к созданию океанических зональных шкал этого интервала для различных обширных широтных областей: низких широт, южных и северных высоких широт (рис. 17–20). Степень их детальности и обоснованности неодинакова, что обусловлено разным количеством данных по смене комплексов в непрерывных полных разрезах, а также недостатком прямых корреляций с палеомагнитной шкалой и зонами по карбонатному микропланктону. К настоящему времени прямые корреляции с палеомагнитной шкалой выполнены только для олигоценовых зон высоких южных и низких широт. К недостаткам схемы для Норвежско-Гренландского бассейна (рис. 20), где изучены представительные палеогеновые диатомовые комплексы, но разрезы охарактеризованы диатомеями неполностью и имеют ряд перерывов, относится несмыкаемость ряда биостратиграфических подразделений.
Рис. 17. Зональная океаническая шкала среднего эоцена – олигоцена низких широт по диатомеям и границы стратиграфического распространения маркирующих видов
Рис. 18. Зональная океаническая шкала среднего эоцена–олигоцена южных высоких широт по диатомеям и границы стратиграфического распространения маркирующих видов (из Scherer et al., 2007)
Рис. 19. Зональная шкала по диатомеям олигоцена – нижнего миоцена Северной Пацифики (по Gladenkov, Barron, 1995; Гладенков, 2007)
Рис. 20. Зональные подразделения по диатомеям для расчленения среднего эоцена–олигоцена
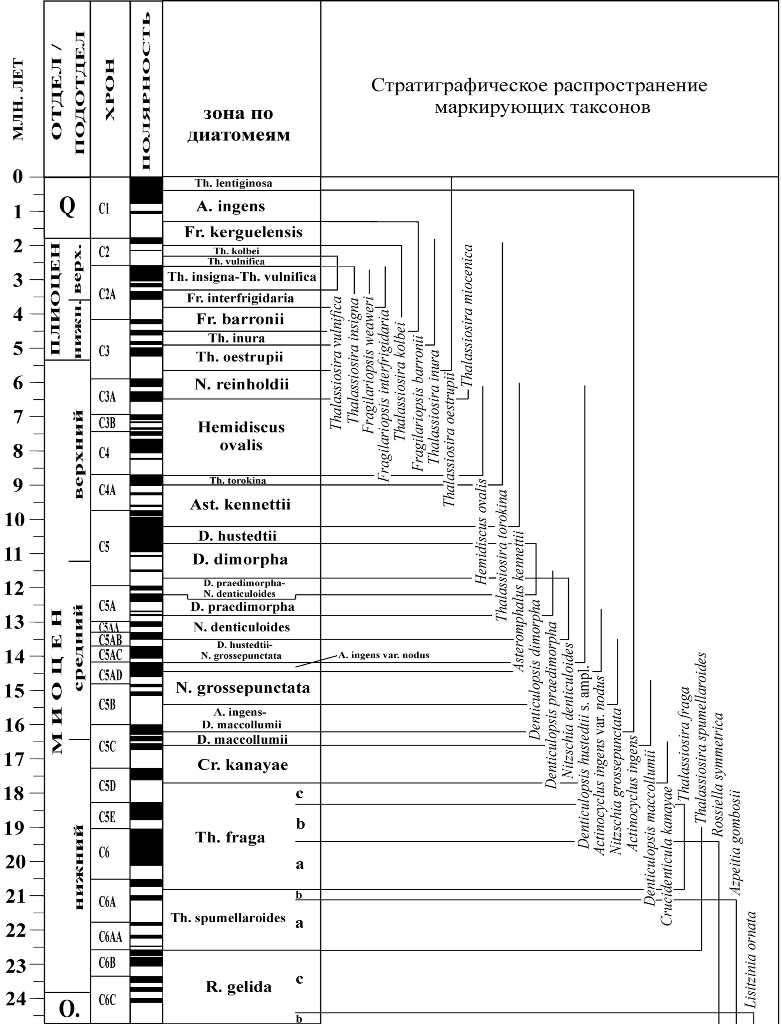
Несмотря на определенное несовершенство зональных шкал, разработанных, прежде всего по материалам глубоководного бурения в океанах, их создание имеет огромное значение, прежде всего для региональных и межрегиональных корреляций. Например, это сделало возможным использование при работе с разрезами суши общих с океаном форм, с установленными интервалами стратиграфического распространения. В целом, такой анализ показал необходимость пересмотра возраста и объема многих выделенных ранее подразделений, относившихся к зонам. Поэтому, например, были предложены новые варианты зональных региональных схем для расчленения и корреляции палеогена на европейской территории бывшего СССР, в которых были использованы сопоставления с океаническими шкалами. 8.5. Зональные шкалы неогена и квартера Зональные схемы расчленения морского неогена и квартера отличаются большей детальностью по сравнению с палеогеном. Остатки диатомовых широко представлены в разрезах верхнего кайнозоя практически во всех районах Земного шара. Изучение большого количества материалов, полученных, в первую очередь, при глубоководном бурении, позволило достаточно детально проследить смену ископаемых ассоциаций в непрерывных разрезах и на этом основании предложить дробные стратиграфические схемы. В настоящее время для расчленения морских отложений нескольких крупных областей (низкие широты, Южный океан, средние-высокие широты Тихого океана) разработаны зональные шкалы по диатомеям, отличающиеся по степени дробности (рис. 21–23). Их зональные комплексы отличаются по составу, а поэтому характеристики зональных границ основаны на фиксировании последовательности разных датированных уровней. В качестве биостратиграфических маркеров в шкалах используются различные виды, типичные для данной обширной области и имеющие в ее пределах устойчивые границы стратиграфического распространения. Зоны обычно имеют широкорегиональный масштаб, хотя определенный провинциализм этой группы сказывается на представительности зон в разных районах. Границы биостратиграфических зональных и внутризональных подразделений в большинстве случаев напрямую скоррелированы с магнитостратиграфической шкалой, что позволяет реально оценивать их возраст.
Рис. 21. Океаническая зональная шкала по диатомеям для расчленения неогена–квартера* низких широт (по Barron, 2003, 2005; Barron et al., 2004)
Рис. 22. Океаническая зональная шкала по диатомеям для расчленения неогена-квартера высоких южных широт, сопоставленная с палеомагнитной шкалой (по Harwood, Maruyama, 1992; Barron, 2003)
Рис. 23. Океанические зональные шкалы по диатомеям для расчленения неогена–квартера Южного океана (А), низких широт (Б) и Северной Пацифики (В) (по Scherer et al., 2007)
8.6. Применение зональных подразделений В настоящее время диатомовый анализ активно используется в геологической практике. Особенно большое значение изучение диатомей имеет при расчленении осадочных толщ олигоцена–квартера областей высоких и средних широт, так как остатки карбонатного микропланктона в них отсутствуют или очень редки. Разработанные зональные океанические шкалы используются не только при исследовании осадочных отложений, формировавшихся в пределах открытого океана. Они также успешно применяются при датировании, расчленении и корреляции толщ окраиноморских и наземных разрезов. В последние десятилетия сопоставление найденных в таких разрезах диатомовых комплексов с ассоциациями зональных шкал позволило во многих случаях провести дробное расчленение кайнозойских толщ (в том числе, считавшихся «немыми»), а также уточнить или пересмотреть возраст многих свит, горизонтов и формаций. Необходимо, однако, отметить, что глобальных верхнекайнозойских зон по диатомеям в строгом смысле не существует, так как диатомовые комплексы, по крайней мере, с олигоцена, характеризуются определенной степенью провинциализма и приурочены к определенным широтным поясам. С учетом того, что все эти регионы очень обширны, можно относить выделенные в них зональные подразделения (составляющие последовательность смыкающихся биозон с устойчивыми границами) к зонам широкорегионального масштаба. Такие зоны прослеживаются в пределах значительных сегментов океана и его обрамления, охватывающих по площади тысячи квадратных километров. Что касается пресноводных диатомей, то при исследованиях континентальных (прежде всего озерных) отложений, они также представляют достаточно эффективный биостратиграфический инструмент. В этом отношении ярким примером последних лет служат результаты, полученные при исследовании осадочных толщ озера Байкал. Изучение ископаемой флоры из разрезов скважин, пробуренных в Байкале, позволило на детальной (зональной) основе расчленить отложения верхнего миоцена – голоцена и с помощью физических методов оценить продолжительность выделенных зон (Кузьмин и др., 2009). При работе с пресноводными древними диатомеями обычно выделяются локальные или местные биостратиграфические подразделения. В отличие от зон по морским диатомовым они используются при расчленении и корреляции отложений в пределах достаточно ограниченных площадей – территорий, к которым был приурочен древний водоем или озерная система. Это объясняется значительной степенью провинциализма флор континентальных бассейнов, а также различными пределами стратиграфического распространения маркирующих видов.
9. Основные черты эволюции диатомей Описанные выше климатические и географические изменения в истории Земли определили три взаимосвязанных, но принципиально разных явления в эволюции диатомовых водорослей: филогенез, флорогенез и филоценогенез. Филогенез предполагает изучение родственных связей между таксонами и выстраивание филогенетических древ. В настоящее время есть публикации по отдельным таксонам диатомовых водорослей, но единой картины филогенеза диатомей не предложено. Молекулярно-генетические исследования позволяют выявлять родственные группы, но для вымерших форм это невозможно. Система филоценогенеза предполагает изучение эволюции (изменения во времени) ценозов, пока также не разработана для диатомей (если не принимать во внимание историческую смену комплексов диатомей). Лучше изучены закономерности флорогенеза морских диатомовых водорослей. Флора – это исторически сложившаяся совокупность видов, или таксонов иного уровня. Формирование флоры – флорогенез – основано на процессах новообразования, миграции видов и вымирании, которые изменяют состав и соотношение видов, родов и таксонов более высокого уровня. Наиболее древние диатомеи найдены в раннемеловых (апт-альбских) отложениях. Изучение остатков диатомей показало, что их таксономический состав уже достаточно разнообразен, а представители различных таксонов имеют панцири, отличающиеся по строению и морфологическим особенностям. Поэтому не исключено, что диатомовые водоросли могли появиться в более древнюю эпоху, но по разным причинам не сохраниться в геологической летописи. Строение и морфология панцирей древнейших (центрических) форм, а также наличие спор могут указывать на их преимущественно бентосный образ жизни в мелководных бассейнах (Harwood et al., 2007). Несколько позже в раннемеловое время появляются первые планктонные диатомеи. Первая зафиксированная стадия развития диатомей заканчивается в начале позднего мела, когда вблизи границы между сеноманом и туроном происходит вымирание большинства существовавших таксонов, относящихся всего к нескольким порядкам (Harwood, Nikolaev, 1995; Harwood et al., 2007). С позднемеловой эпохи начинается активное эволюционное развитие диатомей, с проявлением большой степени разнообразия и появлением многих порядков, давших начало современной флоре. С этого времени они получают достаточно широкое распространение в различных морских зонах, где начинают отличаться высокой степенью продуктивности и приобретать породообразующее значение, о чем свидетельствуют наиболее древние (позднемеловые) диатомиты. Несмотря на имеющиеся особенности флор в разных географических зонах, намечается единая принципиальная картина последовательности этапов флорогенеза. Общую направленность флорогенеза можно рассматривать как отражение естественной этапности эволюционного развития диатомовых водорослей (Стрельникова, 1992). Позднемеловая и раннепалеоценовая флоры диатомовых характеризуются преимущественным развитием представителей порядка Biddulphiales. Массового вымирания диатомей на родовом и видовом уровнях на границе между мелом и палеогеном не происходило. Более 85% видов продолжают встречаться и в палеоцене. Наиболее крупная перестройка флоры произошла на рубеже между поздним палеоценом и ранним эоценом и связана с достаточно хорошо изученным событием PETM (Paleocene-Eocene Therminal Maximum – палеоцен-эоценовый термический максимум) (Barron et al., 2015). Палеоценовая флора с доминирующем порядком Biddulphiales завершает меловой этап в развитии диатомовых водорослей. Следующий эволюционный этап охватывает флоры эоцена и раннего олигоцена и характеризуется сменой биддульфиоидных форм косцинодискоидными и расцветом порядка Coscinodiscales. Максимум в развитии флор этого этапа приходится на средний эоцен. В среднеэоценовое время наблюдается максимальное разнообразие и богатство флоры диатомовых палеогена. Следующий эволюционный этап развития флор начался в позднеолигоценовое время. Для морских флор этого этапа (который, в целом, продолжается до настоящего времени) характерно интенсивное формирование порядка Thalassiosirales и пеннатных диатомей. У представителей флоры этого типа формируются новые морфологические структуры, например, выросты с опорами (фультопортулы). Начало формирования диатомовых флор фитогеографических областей (Аркто-бореальной, Тропической, Антарктической), существующих в современном Мировом океане, по-видимому, относится к среднему эоцену, когда тропическая флора достигает высокой степени эндемизма на уровне родов и видов. В позднеэоценовое и раннеолигоценовое время сохраняется приблизительно такая же географическая дифференциация родов, более резкие отличия отмечаются в позднеолигоценое время. Этапным в развитии диатомей в целом является рубеж вблизи границы между эоценом и олигоценом. С этого времени наблюдается процесс появления и широкого развития таксонов, существующих и в современных водоемах, имеющих створки небольших размеров и с тонкими стенками. Для более же древних, чем олигоценовые, диатомовых ассоциаций характерными, прежде всего, являлись формы с более сильно окремненными и/или большими размерами створок. В олигоцене-неогене диатомовые получили особо широкое распространение в морских экосистемах и начали преобладать в фитопланктоне высоких и средних широт Мирового океана. С этого же времени зафиксировано интенсивное развитие пресноводных диатомей в континентальных водоемах. В целом, имеющиеся к настоящему времени данные свидетельствуют о том, что на протяжении кайнозоя периоды массовых (катастрофических) вымираний морских диатомей отсутствовали. Однако при этом в развитии кайнозойской диатомовой флоры прослеживаются, по крайней мере, шесть периодов относительно быстрых существенных перестроек ассоциаций: 1) вблизи границы между ранним и средним эоценом; 2) в начале раннего олигоцена; 3) вблизи границы между олигоценом и миоценом; 4) в начале среднего миоцена; 5) в конце позднего миоцена; 6) в позднем плиоцене. В эти интервалы происходило постепенное вымирание ряда видов, которые были характерны для составов комплексов в течение миллионов лет, и их замещение новыми видами, появившимися в ходе эволюции. Необходимо также отметить, что к этим интервалам приурочены вымирание и возникновение таксонов не только на видовом и родовом уровнях, но и на уровне семейств и более высоких таксонов.
10. Заключение Диатомовые водоросли представляют очень разнообразную в таксономическом отношении и экологически пластичную группу одноклеточных микроорганизмов, изучение которой представляет большой интерес и важность не только для биологов, но и для палеонтологов, стратиграфов, географов и экологов. Диатомеи являются неотъемлемой составной частью морских и пресноводных экосистем. В современных бассейнах эти водоросли представляют важнейшее звено в цепочке пищевых связей и основных продуцентов органического вещества. Несмотря на то, что диатомовые населяют только эвфотическую зону (то есть зону фотосинтеза, самую верхнюю «пленку» вод максимально до 150 м глубиной), они составляют по биомассе бóльшую часть массы всех водорослей и поставляют около 50% органического вещества. В то же время диатомеи являются основными продуцентами биогенного кремнезема и главными его поставщиками в донные осадки при седиментации. Достоверно диатомовые водоросли найдены в раннемеловых отложениях, однако более широкое распространение диатомей зафиксировано с эоцена, а в олигоцене-неогене они начали преобладать в фитопланктоне Мирового океана, достигнув в среднем миоцене-плиоцене максимальной степени развития и продуктивности (на что указывают самые высокие темпы накопления биогенных кремнистых осадков и масштабы площадного распространения таких отложений). Диатомеи служат достаточно действенным инструментом при детальном стратиграфическом расчленении и определении возраста толщ кайнозоя (прежде всего, морских). Разработанные по этой группе зональные шкалы находят широкое применение в современной геологической практике. С другой стороны, результаты, полученные при исследованиях таксономического состава, закономерностей пространственного распространения и продуктивности сообществ диатомей, направленности их развития, представляют ценный материал для проведения палеоклиматических, палеогеографических и палеоокеанологических реконструкций. Выполненные на базисе детальной стратиграфии построения позволяют определять возраст различных биотических и абиотических событий прошлого, а также подходить к их анализу, оценке и возможной связи.
[1] Более подробно о воспроизведении диатомовых водорослей см. в статье Н.А. Давидовича (2017). [2] Более подробно о морфологии диатомовых водорослей и морфологических терминах см. в статье Р.М. Гогорева и др. (2018).
Авторы заявляют об отсутствии конфликта интересов, требующего раскрытия в данной статье.
Список литературы
Рекомендуемая литература
Статья поступила в редакцию 06.03.2019
Об авторах Стрельникова Нина Ивановна – Nina I. Strelnikova доктор биологических наук, профессор diatomspb@yandex.ru Гладенков Андрей Юрьевич – Andrey Yu. Gladenkov доктор геолого-минералогических наук agladenkov@ilran.ru Корреспондентский адрес: 119017, Россия, г. Москва, Пыжевский пер., 7, Геологический институт РАН. Телефон (495)951-27-23.
ССЫЛКА: Стрельникова Н.И., Гладенков А.Ю. Диатомовые водоросли и их использование в стратиграфических и палеогеографических исследованиях // Вопросы современной альгологии. 2019. № 2 (20). С. 1–38. URL: http://algology.ru/1486 DOI – https://doi.org/10.33624/2311-0147-2019-2(20)-1-38
При перепечатке ссылка на сайт обязательна
Diatoms and their application in stratigraphic and paleogeographic studies Nina I. Strelnikova1, Andrey Yu. Gladenkov2 1Saint Petersburg State University (Saint Petersburg, Russia)
Modern data on diatom algae – unicellular microorganisms with siliceous frustule composing Class Bacillariophyceae, are reviewed. Brief characteristic of biology, morphology, frustule structure and its elements, principles of diatom systematics, and also biogeography and ecology are given. A role of diatoms in sedimentation and peculiarities of preservation of their remains in bottom sediments are discussed. Special attention is paid to issues of effective application of data on fossil diatoms for detailed biostratigraphic subdivision, dating and correlation of sedimentary successions (first of all, marine deposits) as well as for paleoenvironmental reconstructions. General characteristic of diatom zonations proposed for marine Cretaceous and Cenozoic sediments is given. Main features of diatom evolution are discussed. Key words: diatoms; biology; morphology; frustule structure; ecology; biogeography; biosiliceous sedimentation; application in biostratigraphy and paleoenvironmental reconstructions; general features of evolution.
References
Suggested reading
Authors Strelnikova Nina I. Saint Petersburg State University, Saint Petersburg, Russia diatomspb@yandex.ru Gladenkov Andrey Yu. ORCID – https://orcid.org/0000-0002-3378-2155, eLIBRARY Author ID – 61439. Geological Institute of RAS, Moscow, Russia agladenkov@ilran.ru
ARTICLE LINK: Strelnikova N.I., Gladenkov A.Yu. Diatoms and their application in stratigraphic and paleogeographic studies. Voprosy sovremennoi algologii (Issues of modern algology). 2019. № 2 (20). P. 1–38. URL: http://algology.ru/1486 DOI – https://doi.org/10.33624/2311-0147-2019-2(20)-1-38 When reprinting a link to the site is required
На ГЛАВНУЮ

|
|||
|
| ||