|
|
|
|
с международным участием «Водоросли: проблемы таксономии, экологии и использование в мониторинге», посвященной памяти Веры Ивановны Есыревой (Нижний Новгород) Соотношение между биомассой и площадью поверхности харовых водорослей
Макаревич Т.А., Борейко Н.В. Tamara A. Makarevich, Natalia V. Boreiko
Белорусский государственный университет (Минск, Беларусь)
УДК 582.263.3
Исследована зависимость между площадью поверхности и биомассой харовых водорослей в олиго-мезотрофном озере Нарочь (Беларусь). Площадь поверхности определена методом геометрического подобия после разделения таллома на отдельные фрагменты цилиндрической формы. Оценены величины удельной поверхности (S/W) харовых водорослей в разные сезоны года и на различных глубинах. Сезонные изменения удельной поверхности выражены слабо. Средняя за период наблюдения (июль 2018 – март 2020 гг.) величина S/W составила 194,3±29,6 см2/г абсолютно сухой массы (cv=15%). Удельная поверхность харовых водорослей несколько увеличивается с возрастанием глубины. Между массой и площадью поверхности харовых водорослей существует положительная связь, которая удовлетворительно описывается уравнением линейной регрессии: y=130,6x+134,1 (y – площадь поверхности, см2; x – абсолютно сухая масса, г; R2=0,41; P<0,000001). Уравнение может быть использовано для оценки площади поверхности харовых водорослей через сравнительно легко определяемую величину биомассы при масштабных исследованиях, или в случае экспертных оценок, когда не нужна высокая точность. Ключевые слова: харовые водоросли; площадь поверхности; биомасса; удельная поверхность; озеро Нарочь
Введение Харовые водоросли доминируют среди погруженных макрофитов во многих озерах. Примером может служить озеро Нарочь в Беларуси, в котором зона зарослей погруженных макрофитов составляет до 30% площади ложа, а на долю харовых водорослей приходится около 75% площади подводных зарослей и почти 80% биомассы погруженных макрофитов (Экосистема Нарочанских озер, 1985). Оценка роли в экосистемных процессах харовых водорослей (как и макрофитов в целом) базируется, как правило, на определении биомассы. Оценки поверхности харовых водорослей, таллом которых рассечен на мелкие доли, в силу трудоемкости крайне редки. Значимость этого параметра определяется двумя моментами. Во-первых, поверхность макроводорослей, равно как и одноклеточных микроводорослей, представляет собой биоконтур, через который осуществляются обменные процессы с окружающей средой. Известна зависимость интенсивности внешнего массообмена и продукции макро- и микроводорослей от их удельной поверхности (Завалко и др., 1993; Завалко, Шошина, 2008; Хайлов, Парчевский, 1983; Хайлов и др., 1992; Щур и др., 1998). Во-вторых, харовые водоросли – удобный субстрат для перифитона. Интегральная поверхность харовых водорослей в озере – составная часть перифитали, а в случае их доминирования, как в оз. Нарочь, основная. От размера перифитали, т.е. жизненного пространства для прикрепленных сообществ, зависит роль перифитона в продукционно-деструкционных процессах, в формировании биоразнообразия экосистемы. Кроме того, определение величины поверхности харовых водорослей необходимо для корректной оценки плотности и биомассы перифитона на них. Ввиду трудности определения площади поверхности не только харовых водорослей, но и большинства высших водных растений, количество перифитона чаще всего выражают на единицу биомассы макрофитов. Такие величины не могут быть использованы для сравнения обилия перифитона на разных видах макрофитов, поскольку, согласно имеющимся данным, величины удельной поверхности макрофитов (S/W) могут существенно различаться (Макаревич и др., 2019; Armstrong et al., 2003; Sher-Kaul et al., 1995). Например, нами было показано, что S/W для Elodea canadensis Michx. в оз. Нарочь составила 1300–1500 см2/г сухой массы, а для Schoenoplectus lacustris (L.) Palla – всего 60–80 см2/г. Величины массы перифитона в расчете на 1 г сухой массы этих макрофитов различались на порядок (453 и 47 мг/г сухой массы элодеи и камыша соответственно), тогда как в расчете на 1 см2 поверхности их значения были близкими (0,8 и 0,7 мг/см2) (Макаревич, 1983). Для определения площади поверхности макроводорослей и высших водных растений разработан ряд методов, включая калориметрические, планиметрические методы, анализ изображений и др. (Brown, Many, 1985; Cattaneo, Carignan, 1983; Gerber et al., 1994; Sher-Kaul et al., 1995; Миничева, 1987; Миничева и др., 2003). Все эти методы имеют общий недостаток – они сложны методически и трудоемки, поэтому применимы только в специальных исследованиях ограниченного объема. При масштабных исследованиях или в случае экспертных оценок, когда не нужна высокая точность, можно использовать показатель удельной поверхности (S/W). Оценка площади поверхности через сравнительно легко определяемую величину биомассы рассматривается как перспективный подход. К настоящему времени накоплен определенный объем данных, однако их явно недостаточно, к тому же они относятся в основном к высшим водным растениям. Настоящее исследование выполнено с целью установления соотношения между биомассой и площадью поверхности харовых водорослей в озере Нарочь.
Методика исследования Озеро Нарочь – олиго-мезотрофный полимиктический водоем. Площадь водного зеркала составляет 79,6 км2, средняя глубина 8,9 м, максимальная глубина 24,8 м. Дно имеет сложный рельеф, представленный чередующимися впадинами, подводными грядами и платообразными участками. По всему периметру водоема большие площади заняты мелководьями. Озеро характеризуется высокой прозрачностью воды – среднесезонные величины прозрачности в период 2011–2018 гг. составили от 6,0±0,8 до 7,0±1,3 м. Морфометрия озера и высокая прозрачность воды предопределяют обильное развитие макрофитов. Пояс зарослей харовых водорослей достигает глубины 8 м (Экосистема Нарочанских озер, 1985; Бюллетень экологического состояния озер…, 2019). Харовые водоросли (семейство Characeae Gray, 1821) в озере Нарочь представлены восьмью видами – семь представителей рода Chara Linne, 1753, а также Nitellopsis obtusa (Desvaux) J. Gr. Основными ценозообразующими видами являются Chara globularis Thuiller, Ch. ceratophylla Wallr. и Ch. aspera Willd. (Гигевич, 1985). Образцы харовых водорослей для исследования собраны в период с июля 2018 г. по март 2020 г. На мелководье материал собирали вручную, с глубин >1 м – с помощью дайверов, в осенне-зимний период пробы хары отбирали якорем-кошкой. Во всех случаях на каждой станции отбирали значительное количество материала. Собранную хару тщательно перемешивали, затем из общей массы случайным образом отбирали три аликвоты для дальнейшего установления величины удельной поверхности и три аликвоты для определения обводненности и зольности харовых водорослей. Образцы хары освобождали от седиментов, кристаллов карбоната кальция и по возможности от перифитона интенсивным встряхиванием в фильтрованной озерной воде. Очевидно, что прочно прикрепленную фракцию перифитона таким способом удалить невозможно, остается и некоторое количество карбоната кальция на поверхности хары. Удельную поверхность отдельных видов харовых водорослей в данном исследовании мы не определяли, хотя очевидно, что этот вопрос заслуживает внимания. Пробы, как правило, объединяли несколько видов рода Chara. На мелководье преобладала Ch. globularis, с увеличением глубины доля других видов в пробах, прежде всего Ch. aspera, возрастала. Площадь поверхности харовых водорослей определяли методом геометрического подобия, предварительно разделив таллом на отдельные фрагменты цилиндрической формы. Длину фрагментов измеряли миллиметровой линейкой, диаметр и длину мелких фрагментов – с помощью микроскопа (МБС) при 32-кратном увеличении. Площадь поверхности всех фрагментов суммировали. Удельную поверхность (S/W, см2/г) рассчитывали относительно сырой (обсушивание на фильтровальной бумаге до исчезновения капельножидкой воды) и абсолютно сухой (высушивание в сушильном шкафу до постоянного веса при 65°С) массы растений. Сырую массу определяли взвешиванием на электронных весах OHAUS с точностью до 0,01 г, абсолютно сухую – на аналитических весах OHAUS с точностью до 0,0001 г. Зольность харовых водорослей определяли стандартным методом прокаливания в муфельной печи при 450°С до полного удаления органического вещества. Образцы макрофитов предварительно растирали в фарфоровой ступке до порошкообразной массы, тщательно перемешивали и отбирали аликвоты для анализа. Обводненность макрофитов определяли по потерям массы при высушивании до абсолютно сухого веса. Статистический анализ данных проводили при помощи MS Excel.
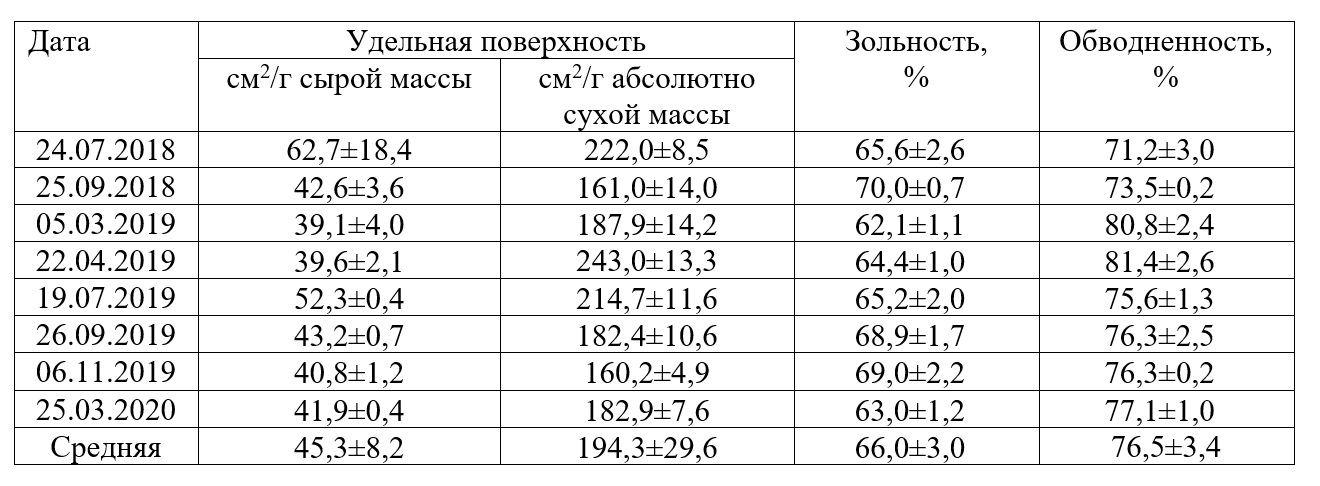
Результаты и обсуждение Имеющиеся в литературе данные указывают на то, что отношение S/W зависит от факторов внешней среды, прежде всего от условий минерального питания растений, освещенности, гидродинамики (Armstrong et al., 2003; Яшкина и др, 2014; Хайлов и др., 1992). Зависит S/W и от возраста макрофита (Завалко, Шошина, 2008). Исходя из этого, мы проанализировали, изменяется ли величина удельной поверхности харовых водорослей в годовом цикле в соответствии с сезонными изменениями комплекса факторов среды. В табл. 1 представлены величины удельной поверхности харовых водорослей в разные сезоны года. Приведены также такие морфофункциональные показатели, как зольность и обводненность, от которых зависит удельная поверхность растений. Данные относятся к мелководной части озера (глубина <1 м). Как правило, растения на мелководье – это организмы одного года вегетации, поскольку во время вскрытия озера хара выносится ледовыми массами на берег. Как следует из полученных результатов, сезонные изменения удельной поверхности харовых водорослей выражены слабо. Средняя за период наблюдения величина S/W в расчете на сырую массу водорослей составила 45,3±8,2 см2/г при коэффициенте вариации 18%, а в расчете на абсолютно сухую массу – 194,3±29,6 см2/г, при коэффициенте вариации 15%. Тем не менее, некоторые тенденции прослеживаются. Более высокие значения S/W наблюдаются в середине вегетационного сезона (июль), более низкие – в подледный период (март). Особенно заметно это проявляется при расчете S/W относительно сырой массы. Летом растения молодые, находятся в стадии активного роста, характеризуются высокой интенсивностью фотосинтеза, идет накопление органического вещества. В подледный период мы наблюдали старые частично отмирающие растения с высокой степенью обводненности таллома. При расчете S/W относительно абсолютно сухой массы прослеживается некоторое снижение показателя осенью. Вероятно, это связано с возрастанием зольности, что увеличивает массу растений. Зольность в осенний период возрастает вследствие накопления в течение вегетационного сезона на поверхности талломов и в межклеточных пространствах карбоната кальция. Харовые водоросли, как известно, являются активными карбонатоосадителями.
Таблица 1. Морфофункциональные показатели харовых водорослей. Table 1. Morphological and functional characteristics of chara algae.
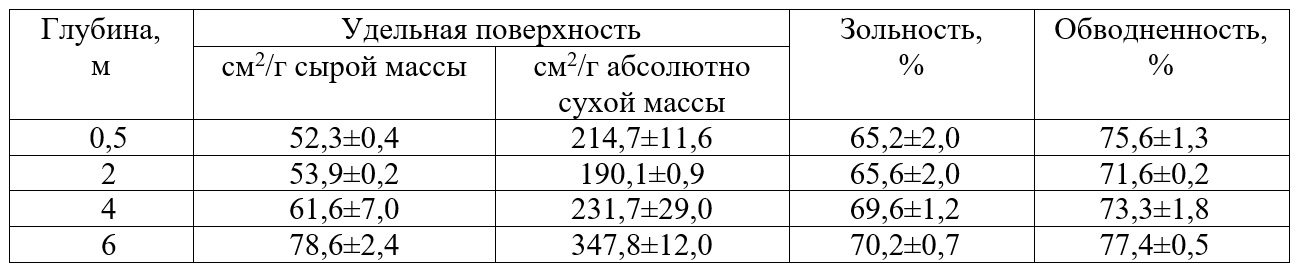
Таблица 2. Морфофункциональные показатели харовых водорослей на разных глубинах оз. Нарочь, 16–19.07.2019 г. (средние значения ± SD; n = 3). Table 2. Morphological and functional characteristics of chara algae at different depths,
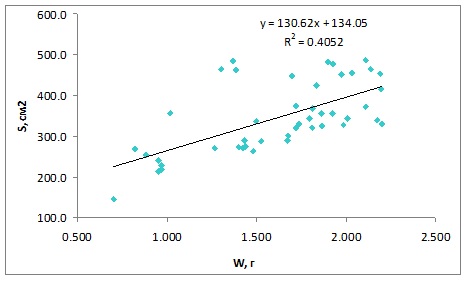
В целом величины S/W харовых водорослей изменяются в достаточно узких пределах. Диапазон величин в расчете на сырую массу составил от 39,1±4,0 до 78,6±2,4 см2/г, а в расчете на абсолютно сухую массу – от 160,2±4,9 до 347,8±12,0 см2/г. Поэтому для расчета математической зависимости между биомассой и площадью поверхности мы объединили все данные, полученные за период исследования. Это позволило увеличить объем выборки. Использовали абсолютно сухую массу водорослей, поскольку сырая масса – достаточно субъективный показатель, определяется с меньшей точностью. Между массой и площадью поверхности харовых водорослей существует положительная связь (рис. 1), которая удовлетворительно описывается уравнением линейной регрессии: y = 130,6x + 134,1 где y – площадь поверхности, см2; Параметры уравнения: свободный член уравнения (±SE) – 134,1±40,4; коэффициент регрессии (±SE) – 130,6±24,1; R2=0,41; P<0,000001.
Рис. 1. Связь между биомассой (W, г абсолютно сухой массы) и площадью поверхности (S, см2) харовых водорослей. Озеро Нарочь, n = 45 Fig. 1. Relationship between biomass (W, g dry weight) and surface area (S, sm2) of chara algae.
Очевидно, что связь между площадью поверхности и массой харовых водорослей, установленная в диапазоне условий среды одного озера, нельзя считать универсальной. Тем не менее, полученные нами результаты указывают на перспективность подхода к оценке площади поверхности харовых водорослей по их биомассе. Методы определения биомассы стандартизированы и сравнительно просты. При проведении масштабных исследований или в случае экспертных оценок, когда не требуется высокая точность, можно использовать данные по биомассе харовых водорослей для оценки их площади поверхности. Авторы заявляют об отсутствии конфликта интересов, требующего раскрытия в данной статье.
Список литературы
Статья поступила в редакцию 20.06.2021
Об авторах Макаревич Тамара Александровна – Tamara A. Makarevich кандидат биологических наук makarta@tut.by Борейко Наталья Витальевна – Natalia V. Boreiko студент boreyko.natali@mail.ru Корреспондентский адрес: 220030, Беларусь, Минск, проспект Независимости, 4, БГУ, Биологический факультет; тел. 375(17)209-58-11.
ССЫЛКА: Макаревич Т.А., Борейко Н.В. Соотношение между биомассой и площадью поверхности харовых водорослей // Вопросы современной альгологии. 2021. №2 (26). С. 8–15. URL: http://www.algology.ru/1694 DOI – https://doi.org/10.33624/2311-0147-2021-2(26)-8-15 При перепечатке ссылка на сайт обязательна Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно.
Relationship between biomass and surface area of Charophyceae Tamara A. Makarevich, Natalia V. Boreiko Belarusian State University (Minsk, Belarus)
Relationships between charophytes surface area and biomass were studied in oligo-mesotrophic Naroch Lake (Belarus). To determine the surface area of Charophytes the geometric similarity method was used, preceded by dividing the thallus into separate cylindrical fragments. The specific surface area (S/W) of chara algae was determined in different seasons and at different depths. Seasonal changes in specific surface area are weakly expressed. The average S/W value for the observation period (July 2018 – March 2020) was (194,3±29,6) sm2/ g dry weight (cv=15%). The specific surface area of charophytes grows slightly with increasing depth. There is a positive relationship between the dry mass and surface area of charophytes, which is satisfactorily described by the linear regression equation: y = 130,6x+134,1 (y – surface area, sm2; x – absolutely dry weight, g; R2=0,41; P<0,00000). The equation can be used to estimate the surface area of chara algae through the relatively easy determination of biomass in large-scale studies, or in the case of expert assessments, when high accuracy is not needed. Key words: Charophyte algae; surface area; biomass; specific surface area; Naroch Lake
References
Authors Makarevich Tamara A. ORCID – https://orcid.org/0000-0003-1437-0187 Belarusian State University, Minsk, Belarus makarta@tut.by Boreiko Natalia V. Belarusian State University, Minsk, Belarus boreyko.natali@mail.ru
ARTICLE LINK: Makarevich T.A., Boreiko N.V. Relationship between biomass and surface area of Charophyceae. Voprosy sovremennoi algologii [Issues of modern algology]. 2021. №2 (26). P. 8–15. URL: http://www.algology.ru/1694 DOI – https://doi.org/10.33624/2311-0147-2021-2(26)-8-15 When reprinting a link to the site is required Dear colleagues! If you want to receive the version of the article in PDF format, write to the editor,please and we send it to you with pleasure for free.
На ГЛАВНУЮ
|
|||
|
| ||