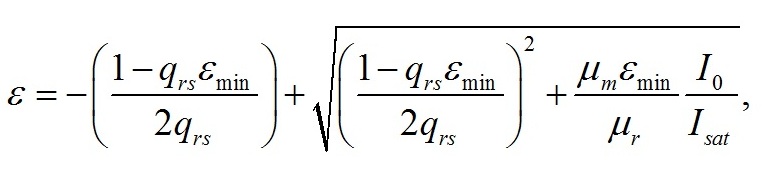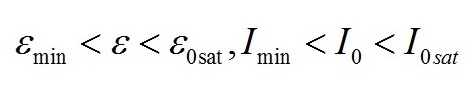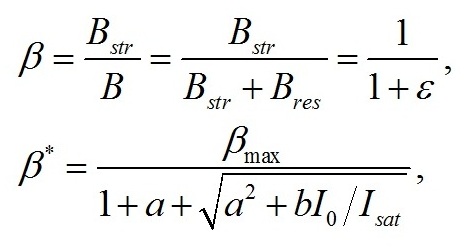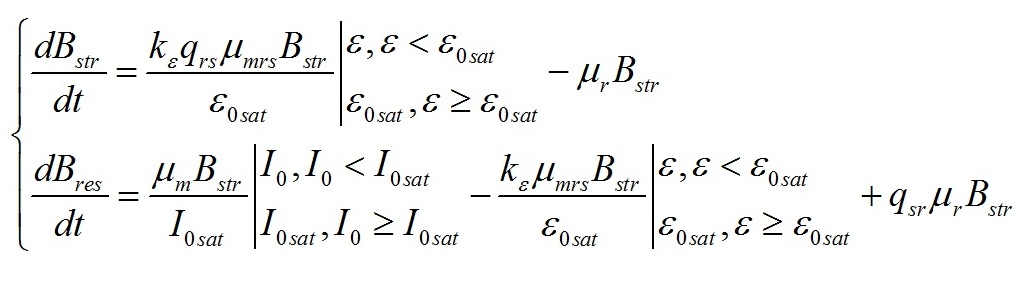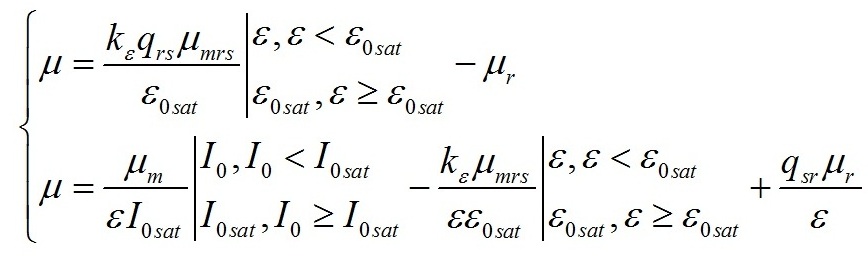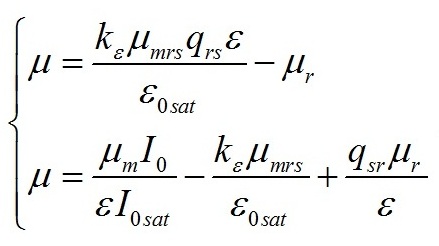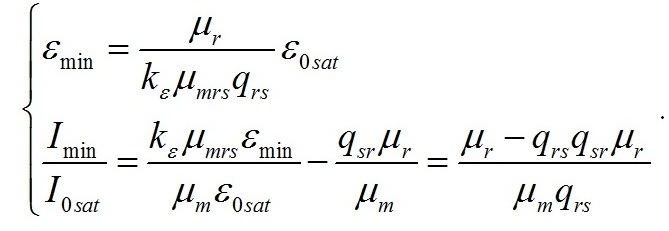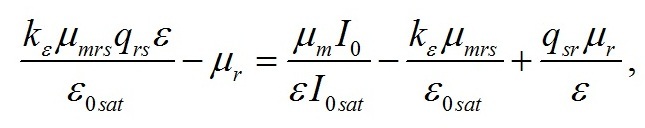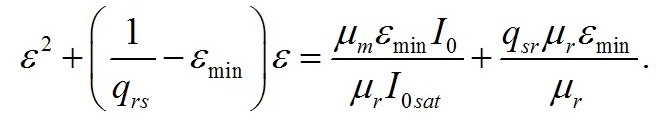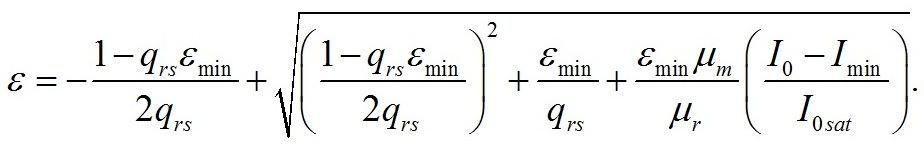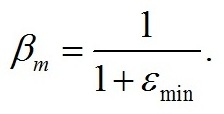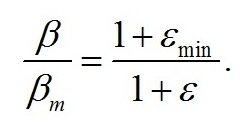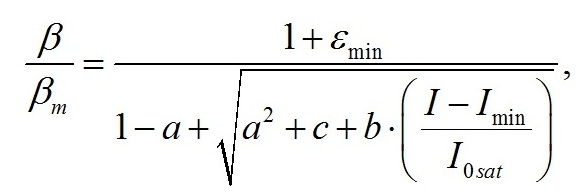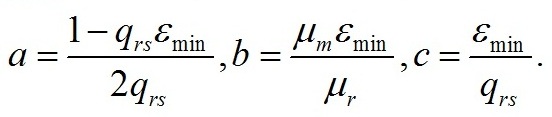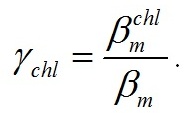|
|
|
|
по Материалам Международной конференции «Экологическая физиология водных фототрофов: распространение, запасы, химический состав и использование» VIII Сабининские чтения 10 ноября 2019 - 29 февраля 2020 Модифицированное уравнение содержания хлорофилла в биомассе микроводорослей при световом лимитировании
Лелеков А. С., Тренкеншу Р.П., Новикова Т.М. Alexander S. Lelekov, Rudolf P. Trenkenshu, Tatyana M. Novikova
Федеральный исследовательский центр
УДК 579.017.8:57.036
В работе представлена модифицированная математическая модель зависимости относительного содержания хлорофилла а в биомассе микроводорослей от облучённости. Основой базовой модели служит представление биомассы как суммы резервных и структурных составляющих. Учитывая, что часть структурной биомассы может превращаться в резервную, в базовое уравнение добавлен параметр минимальной облучённости («компенсационный пункт»). Верификация модели на экспериментальных данных для плотностатных культур Tetraselmis viridis и Arthrospira platensis позволила оценить видоспецифичные коэффициенты. Полученные значения коэффициентов близки для обоих видов. Доля хлорофилла в структурной биомассе составляет около 2%, максимальная доля структурных форм биомассы – 84%, экономический коэффициент перехода резервной биомассы в структурную – 80–90%, максимальная удельная скорость фотосинтеза превышает скорость эндогенного расхода биомассы в 14–20 раз. Ключевые слова: культура микроводорослей; хлорофилл; световые условия; моделирование
Введение При адаптации микроводорослей к различной облучённости происходят изменения пигментного состава клеток. Так, для различных систематических групп микроводорослей с ростом облучённости отмечено уменьшение количества хлорофилла а и других пигментов (Geider et al., 1998; Falkowski, Roche, 1991). Также изменяются соотношения между различными фотосинтетическими пигментами (Richardson et al., 1983). Кроме того, в течение светового периода в клетках микроводорослей происходит синтез и накопление запасных (резервных) углеводов и липидов, которые расходуются в темновой период, в том числе на синтез белков (Torzillo et al., 1991). Качественные модели зависимости относительного содержания хлорофилла а от облучённости базируются на общепринятых представлениях о фотодеструкции пигментов. Несмотря на широкое распространение данного подхода, в литературе можно встретить работы, в которых экспериментально деструкция пигментов не была подтверждена. Так, доля хлорофиллида и других продуктов распада не превышает 2% клеточного содержания хлорофилла а при любых облучённостях (Goericke, Welschmeyer, 1992). Представляется актуальным разработка новых подходов, которые, не отвергая существующих представлений, позволили бы предложить новые базовые принципы моделирования влияния света на фотобиосинтез культур микроводорослей. Предлагаемый нами подход базируется на общепринятой схеме разделения процессов фотобиосинтеза микроводорослей на световую и темновую стадии, а также разделении продуктов этих стадий на резервные и структурные составляющие. Для условий светолимитирования в культурах невысокой плотности соотношение резервных (Bres) и структурных (Bstr) форм биомассы (ε) определяется внешней облучённостью I0 (Тренкеншу, 2017): где εmin – минимальное соотношение резервных и структурных форм биомассы при «компенсационном пункте» фотосинтеза; qrs – экономический коэффициент перехода резервных форм в структурные; μm – максимальная удельная скорость фотосинтеза; μr – удельная скорость эндогенного расхода структурной биомассы. Полученное выражение описывает изменение соотношения ε в зависимости от внешней облучённости при условии отсутствия самозатенения клеток микроводорослей, т. е. при невысоких плотностях культуры. При некоторой облучённости Imin («компенсационный пункт» фотосинтеза), величина ε достигает минимального значения. С ростом интенсивности света ε увеличивается, достигая максимума при I0sat. Таким образом, границы применимости выражения (1): На основе выражения (1) в работе (Тренкеншу, Новикова, 2019) предложена модель зависимости относительного содержания хлорофилла а от внешней облучённости, считая, что пигмент относится к структурной части биомассы: где β*, βmax – относительное и максимальное содержание хлорофилла а в биомассе микроводорослей; а и b – обобщённые параметры. В выражении (3) величина Imin явно не фигурирует, что создает трудности при математических расчётах видоспецифических параметров модели на конкретных экспериментальных данных. Цель данной работы – предложить модифицированное уравнение зависимости относительного содержания хлорофилла а от облучённости, удовлетворяющее граничным условиям (2).
Результаты и обсуждение Общий подход к моделированию фотобиосинтеза и биохимических трансформаций в клетках микроводорослей подробно изложен в работе (Тренкеншу, 2017). В основе (1) лежит система уравнений, которые описывают зависимости скоростей изменения структурной и резервной части биомассы от внешних и внутренних потоков лимитирующего субстрата. В качестве внешнего потока рассматривается поток световой энергии, внутреннего – поток резервных компонентов биомассы на ключевой ферментный комплекс, на котором происходит синтез структурной части. Дополним базовую систему уравнений из (Тренкеншу, 2017) слагаемым, учитывающим то, что часть структурной биомассы может превращаться в резервную. Этот процесс осуществляется в ходе эндогенного катаболизма («темнового дыхания») микроводорослей. Расход части структурной биомассы обеспечивает процессы фотобиосинтеза дополнительным количеством энергии в форме макроэргов, а также «углеродными скелетами» – интермедиатами цикла Кребса (Физиология растений, 2005). Пусть удельная скорость распада структур – μr, эффективность – qsr. Для условий светового лимитирования и культуры невысокой плотности запишем: где qrs – экономический коэффициент перехода резервной биомассы в структурную, I0sat – насыщающая внешняя облучённость и соответствующее её соотношение резервная / структурная биомасса ε0sat, μmrs – максимальная удельная скорость преобразования резервной биомассы в структурную, kε – коэффициент, характеризующий смену лимитирующих факторов, численно равный отношению параметра ε при световом (внешнем) и внутреннем лимитировании. Систему (4) можно записать для удельных скоростей непрерывной культуры: Согласно граничным условиям (2) система уравнений примет вид: Алгоритм нахождения зависимости ε от внешней облучённости I0 аналогичен приведённому в работе (Тренкеншу, 2017). Выразим нижнюю световую границу применимости модели через кинетические параметры культуры микроводорослей. В световом «компенсационном пункте» (I0 = Imin) удельная скорость роста равна нулю, ε = εmin:
Полученные значения минимальных величин (6) используем для нахождения зависимости ε от облучённости. Приравняем первое и второе уравнение системы (5):
Выразим произведение qsr μr из (6) и подставим в последнее уравнение:
Максимальная доля структурных форм (βm) будет наблюдаться при минимальном соотношении резервных и структурных форм биомассы:
Окончательно выражение для β можно записать в виде:
где a, b,c – обобщённые коэффициенты:
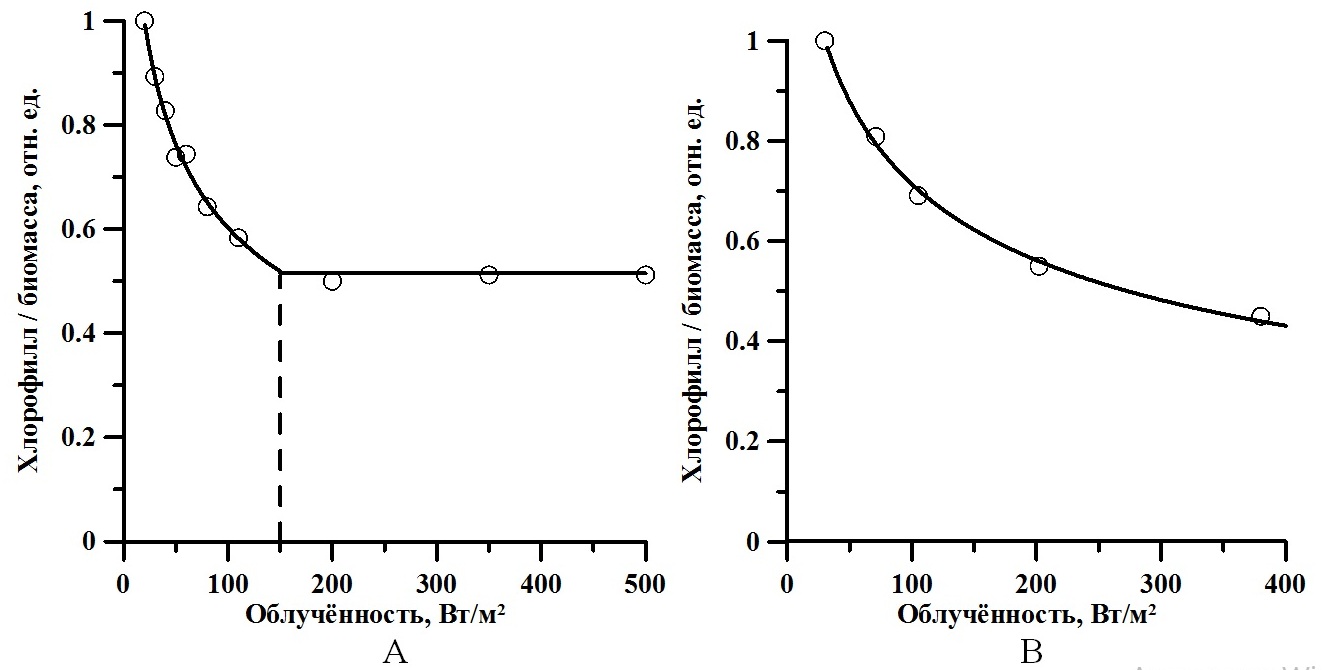
Верификация модели В качестве примера применения (8) рассмотрим экспериментальные данные зависимости относительного содержания хлорофилла а в биомассе Tetraselmis (Platymonas) viridis (по данным Терсков и др., 1979) и Arthrospira (Spirulina) platensis (по данным Заворуева, 2007) от облучённости. Значения точек согласно (8) даны в относительных единицах (каждое значение нормировано на максимальное). Для уменьшения количества неизвестных параметров численные значения компенсационной и насыщающей облучённости были заданы: для T. viridis – 20 и 150 Вт·м-2, для A. platensis – 30 и 380 Вт·м-2 соответственно. Данные значения следует рассматривать как примерные, их величины могут быть уточнены при большем количестве экспериментальных точек. Для T. viridis (см. рис. 1А) в диапазоне интенсивностей света выше насыщающей (150 до 500 Вт/м2) содержание хлорофилла в биомассе не изменялось.
Рис. 1. Зависимость относительного содержания хлорофилла а в биомассе T. viridis (A) (по данным Тренкеншу, 1984) и A. platensis (В) (по данным Заворуева, 2007) от облучённости. Сплошная линия – аппроксимация данных выражением (8) (R2=0,99) Fig. 1. Dependence of the relative chlorophyll a content in T. viridis (A) (according to Trenkenshu, 1984) and A. platensis (B) (according to Zavorueva, 2007) on irradiation. Solid line shows data approximation with equation (8) (R2=0.99)
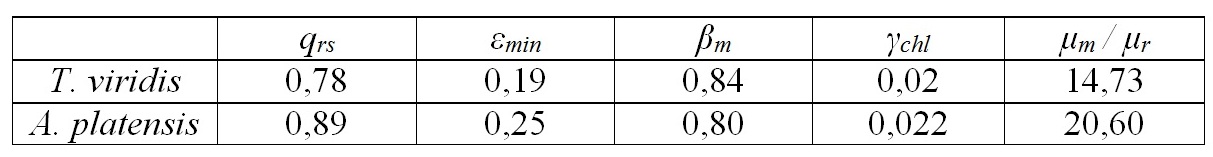
Аппроксимация экспериментальных данных моделью (8) представлена на рис. 1. Определены обобщённые коэффициенты модели, которые позволили рассчитать видоспецифические параметры культур. Так, для двух указанных видов отношение скорости эндогенного расхода биомассы к максимальной удельной скорости фотосинтеза составляет 0,05–0,07. Данная величина варьирует в диапазоне 0,05–0,65 для микроводорослей различных систематических групп при различных условиях культивирования (Geider, Osborne, 1989). Экономический коэффициент перехода резервной биомассы в структурную qrs составил около 80–90%, минимальное соотношение резервная / структурная биомасса – 20–25% (см. табл. 1). Определим долю хлорофилла а в структурной биомассе. Для этого разделим максимальные экспериментальные значения относительного содержания пигмента на рассчитанное максимальное содержание структурных форм:
Полученные значения γchl для двух исследуемых видов близки и составляют около 2%. Это ещё раз подтверждает, что хлорофилл а относится к структурным формам биомассы. Ранее нами для этих же видов показана прямая пропорциональная зависимость между концентрацией общего белка и хлорофилла а (Тренкеншу и др., 2018).
Таблица 1. Видоспецифичные параметры модели (8) для T. viridis и A. platensis Table 1. The species specific parameters of the model (8) to T. viridis and A. platensis
Заключение Предложенный модельный подход, основанный на общих представлениях о процессах фотобиосинтеза культур микроводорослей, позволяет объяснить снижение относительного содержания хлорофилла с ростом внешней облучённости. Отметим, что при составлении первичной системы дифференциальных уравнений не учитывалось фотодеструктивное окисление пигмента, а хлорофилл отнесён к структурным составляющим биомассы, при этом его доля в структуре остается неизменной. Получено модифицированное уравнение зависимости относительного содержания хлорофилла а от интенсивности действующего света для культур невысокой плотности. Модель с высокой точностью описывает результаты экспериментальных данных для T. viridis и A. platensis. Рассчитанные параметры модели необходимо рассматривать как оценочные, значения которых могут уточняться при верификации (8) на большом объёме экспериментальных данных. Тем не менее, значения коэффициентов для двух видов микроводорослей близки, что позволяет судить об универсальности предлагаемого подхода.
Авторы заявляют об отсутствии конфликта интересов, требующего раскрытия в данной статье. Работа выполнена в рамках госзадания ФИЦ ИнБЮМ по теме «Исследование механизмов управления продукционными процессами в биотехнологических комплексах с целью разработки научных основ получения биологически активных веществ и технических продуктов морского генезиса» (№ гос. регистрации АААА-А18-118021350003-6).
Список литературы
Статья поступила в редакцию 15.01.2020
Об авторах Лелеков Александр Сергеевич - Lelekov Alexander S. кандидат биологических наук a.lelekov@yandex.ru Тренкеншу Рудольф Павлович – Trenkenshu Rudolf P. кандидат биологических наук r.trenkenshu@rambler.ru Новикова Татьяна Михайловна - Novikova Tatyana M. младший научный сотрудник, Федеральный исследовательский центр «Институт биологии южных морей им. А.О.Ковалевского РАН» (ФИЦ ИнБЮМ РАН), Севастополь, Россия (Kovalevsky Institute of Marine Biological Research RAS, Russia, Sevastopol), Отдел биотехнологий и фиторесурсов nowtanj@yandex.ru Корреспондентский адрес: Россия, 299011, Севастополь, пр. Нахимова, 2, ФИЦ ИнБЮМ РАН; тел. (8692)550795. ССЫЛКА НА СТАТЬЮ: Лелеков А.С., Тренкеншу Р.П., Новикова Т.М. Модифицированное уравнение содержания хлорофилла в биомассе микроводорослей при световом лимитировании // Вопросы современной альгологии. 2020. № 1 (22). С. 17–24. URL: http://algology.ru/1606 DOI - https://doi.org/10.33624/2311-0147-2020-1(22)-17-24
Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно. При перепечатке ссылка на сайт обязательна
Modified equation of chlorophyll content in microalgal biomass under light limitation Alexander S. Lelekov, Rudolf P. Trenkenshu, Tatyana M. Novikova Kovalevsky Institute of Biology of the Southern Seas RAS, Sevastopol, Russia The paper presents a modified mathematical model of light-dependent chlorophyll concentration in the microalgae biomass. The basic model is based on the concept of biomass as a sum of reserve and structural components. Considering that part of the structural biomass can turn into a reserve, the minimum irradiation parameter (“photosynthesis compensation point”) is added to the basic equation. Verification of the model on experimental data for turbidostat cultures of Tetraselmis viridis and Arthrospira platensis allowed us to evaluate species-specific coefficients. The obtained coefficient values are similar for both species. The share of chlorophyll in structural biomass is about 2 %, the maximum share of structural forms of biomass is 84 %, the economic conversion factor of reserve biomass to structural is 80–90 %, and the maximum specific rate of photosynthesis is 14–20 times higher than the rate of endogenous consumption of biomass. Key words: microalgae culture; chlorophyll; light conditions; modeling.
References
Authors Lelekov Alexander S. Orcid ID – https://orcid.org/0000-0003-1213-7963 Kovalevsky Institute of Marine Biological Research of RAS, Sevastopol, Russia a.lelekov@yandex.ru Trenkenshu Rudolf P. Orcid ID – https://orcid.org/0000-0003-3727-303X Kovalevsky Institute of Marine Biological Research RAS, Sevastopol, Russia r.trenkenshu@rambler.ru Novikova Tatyana M. Orcid ID – https://orcid.org/0000-0002-6366-6470 Kovalevsky Institute of Marine Biological Research RAS, Sevastopol, Russia nowtanj@yandex.ru
ARTICLE LINK: Lelekov A.S., Trenkenshu R.P., Novikova T.M. Modified equation of chlorophyll content in microalgal biomass under light limitation.Voprosy sovremennoi algologii (Issues of modern algology). 2020. №1 (22). P. 17–24. URL: http://algology.ru/1606 DOI - https://doi.org/10.33624/2311-0147-2020-1(22)-17-24
Dear colleagues! If you want to receive the version of the article in PDF format, write to the editor,please and we send it to you with pleasure for free. When reprinting a link to the site is required
К разделу ОБЗОРЫ, СТАТЬИ И КРАТКИЕ СООБЩЕНИЯ

|
|||
|
| ||