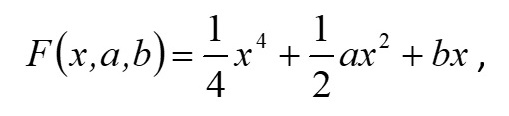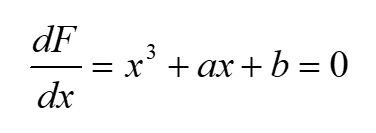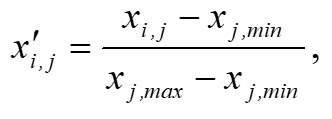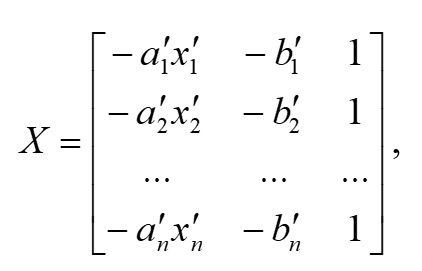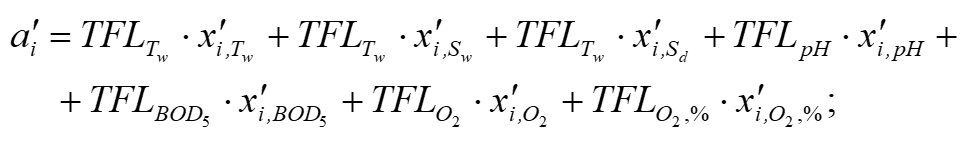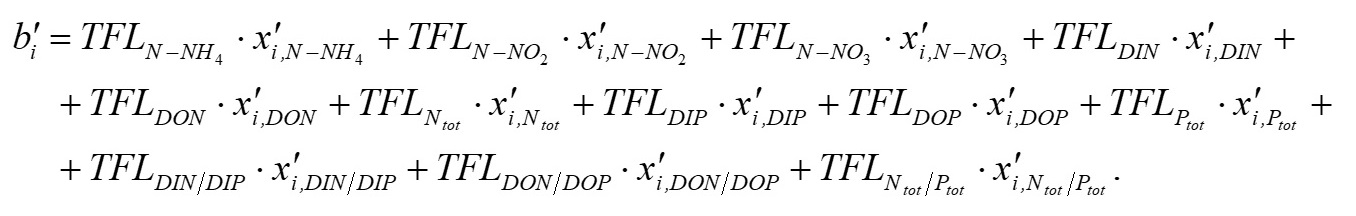|
|
|
|
с международным участием «Водоросли: проблемы таксономии и экологии, использование в мониторинге и биотехнологии» (12–18 сентября 2022, Москва, Россия) Опыт использования методов теории катастроф при описании динамики фитопланктона в Вислинском заливе Балтийского моря
Konstantin A. Podgornyy, Olga A. Dmitrieva 1Атлантический филиал ФГБНУ «ВНИРО» – «АтлантНИРО» (Калининград, Россия) УДК 574.58.001.573+519.6
Для анализа «режимных сдвигов» в экосистемах достаточно широко применяют идеи и методы теории катастроф. В данной работе анализ состояния фитопланктона российской части Вислинского залива Балтийского моря выполнен на основе использования наиболее простой из возможных моделей теории катастроф – катастрофы сборки. В качестве переменной состояния рассмотрена суммарная биомасса фитопланктона. Расчет значений управляющих параметров выполнен с использованием факторных нагрузок главных компонент для данных, которые характеризуют гидрологический режим и уровень биогенной нагрузки на водную экосистему залива. Было установлено, что в зависимости от конкретной ситуации один и тот же фактор или разные комбинации факторов могут играть как стабилизирующую роль, так и быть причиной потери устойчивости. Ключевые слова: водные экосистемы; интегрированный анализ; математическое моделирование; режимный сдвиг; теория катастроф; катастрофа сборки; устойчивость экосистем
Введение Возможность существования резких (часто внезапных), значительных по силе и различных по своей продолжительности динамических изменений в водных экосистемах различного типа представляет собой достаточно известный факт (Scheffer et al., 1993; Scheffer, Carpenter, 2003; Dahlgren, Kautsky, 2004; Folke et al., 2004; Munkes, 2005; Van Nes, Scheffer, 2007; Viaroli et al., 2008; Schallenberg, Sorrell, 2009; Rosqvist et al., 2010; Kornijów, 2018; Thresholds Database – https://www.resalliance.org/thresholds-db). В настоящее время исследователи-экологи нередко расходятся во мнениях относительно тех механизмов, которые приводят к появлению режимных сдвигов и существенных перестроек пространственно-временной структуры экосистем (Petraitis, Dudgeon, 2016). Для анализа такого рода процессов достаточно широко используют идеи и методы теории катастроф (Постон, Стюарт, 1980; Гилмор, 1984; Арнольд, 2004; Том, 2006). В ней проводится качественный анализ сложных нелинейных явлений и поведения динамических систем. Элементарная теория катастроф сводит огромное многообразие ситуаций к небольшому числу стандартных схем, которые можно детально исследовать. Анализ качественного поведения нелинейных динамических систем при изменении описывающих их управляющих параметров позволяет изучать состояния, далёкие от равновесия, а также предсказывать резкую смену этих состояний. Теория катастроф представляет своего рода программу прогнозирования неустойчивости различных по своей природе систем, в том числе и экосистем. Такое название она получила потому, что потеря устойчивости может быть достаточно катастрофична даже если она не приводит к разрушению и/или гибели системы, а лишь только обусловливает ее переход к иной траектории развития. Достаточно часто в теории динамических систем рассматривают режимный сдвиг, который описывают такой известной моделью, как «катастрофа сборки» (сusp catastrophe). Эта модель имеет всего лишь одну обобщенную переменную состояния (так называемый «потенциал», или «потенциальную функцию») и два управляющих параметра (Постон, Стюарт, 1980; Гилмор, 1984; Арнольд, 2004). Для катастрофы типа «сборка» существуют области параметрического пространства (в данном случае это наборы параметров, которые описывают условия окружающей среды), в которых может иметь место только одна устойчивая точка равновесия. Вместе с тем имеются также и такие области, в пределах которых существуют не одна, а три точки равновесия, две из которых устойчивые, а одна – неустойчивая. В зависимости от изменения значений управляющих параметров система может находиться в разных состояниях, а также демонстрировать сложную временную динамику и внезапно «перескакивать» между устойчивыми состояниями. Концепция альтернативных стабильных (устойчивых) состояний находит все более широкое применение при описании и анализе процессов в водных экосистемах. Однако попытки реализовать данную концепцию для объяснения процессов, которые наблюдают в устьях рек и в лагунах, в частности, лагунах Балтийского моря, редки (Dahlgren, Kautsky, 2004; Munkes, 2005; Rosqvist et al., 2010; Kornijów, 2018). Прибрежные лагуны по самой своей природе нестабильны. Переход от континентальных условий к морским неизменно сопровождается появлением седиментологических, гидрологических и биологических градиентов (Viaroli et al., 2008). Все это делает лагуну сложной мозаикой мест обитания. Кроме того, влияние природных и антропогенных факторов может вызывать широкую пространственно-временную изменчивость процессов, что приводит к существенным изменениям экосистемы и связанных с ней структуры сообществ. В рамках данной работы рассмотрены следующие взаимосвязанные задачи:
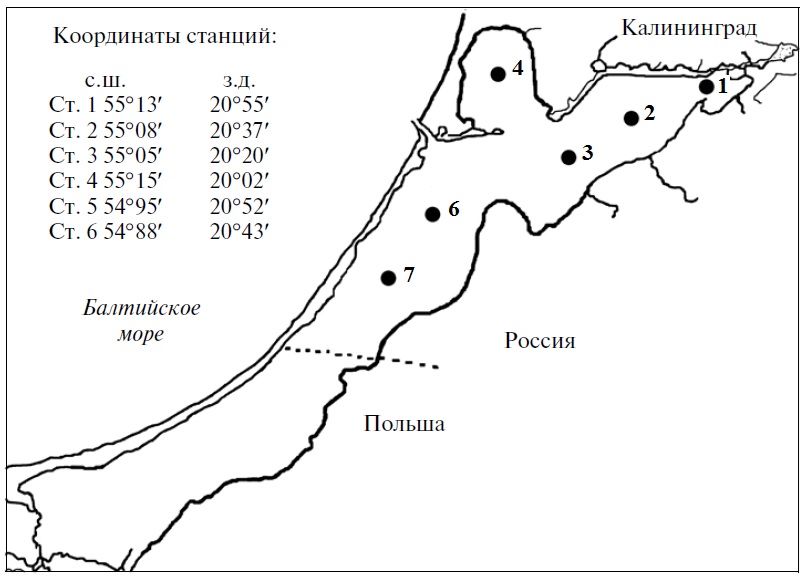
Материалы и методы Вислинский залив расположен в юго-восточной части побережья Балтийского моря и представляет собой узкую, вытянутую вдоль берега лагуну (рис. 1). От моря залив отделен песчаной косой и соединен с ним проливом. Гидрологический режим Вислинского залива определяется метеорологическими условиями, водообменом с Балтийским морем, незначительным поступлением речных вод и его мелководностью. Для залива в целом характерна гомотермия, которая обусловлена мелководностью залива и достаточно частыми и интенсивными процессами ветрового перемешивания. Прозрачность воды залива вследствие небольших его глубин, частых волнений, взмучивающих донные осадки, и высокого содержания органической и неорганической взвеси, довольно небольшая. Ежегодно в Вислинский залив поступают большие объемы различных форм азота и фосфора. Залив является аккумулятором осадочного вещества, в том числе и загрязняющих веществ. Вислинский залив можно классифицировать как опресненный морской водоем. Среднемноголетняя соленость в заливе – 3,7‰. В современный период времени Вислинский залив можно характеризовать как эвтрофный водоем. Гидрохимический режим экосистемы Вислинского залива во многом определяется особенностями его гидрологического режима. Существенное влияние оказывает расположение залива и его водосборной площади в густонаселенном районе с интенсивно развитой промышленностью, сельским хозяйством и судоходством. Исследования фитопланктона в пределах российской части акватории Вислинского залива проводили ежемесячно (с апреля по ноябрь) в рамках программы многолетнего мониторинга ФГБНУ «АтлантНИРО». Пробы фитопланктона отбирали в 2002–2010 гг. на шести стандартных станциях, расположение которых показано на рис. 1. Пробы фитопланктона фиксировали раствором Усачева и сгущали седиментационным методом. Обработку проб проводили в камере «Учинская» (с помощью микроскопа Микромед-2). Биомассу фитопланктона определяли исходя из оценки численности каждого таксона, объема клеток его особей и приравнивая удельную массу водорослей к 1. Объемы клеток, соответствующие геометрическим фигурам (шару, цилиндру и двум сопряженным конусам), вычисляли с помощью таблиц для перевода линейных размеров в объемные.
Рис. 1. Станции отбора проб фитопланктона Fig. 1. Phytoplankton sampling stations in the Kaliningrad (Vistula) Lagoon of the Baltic Sea
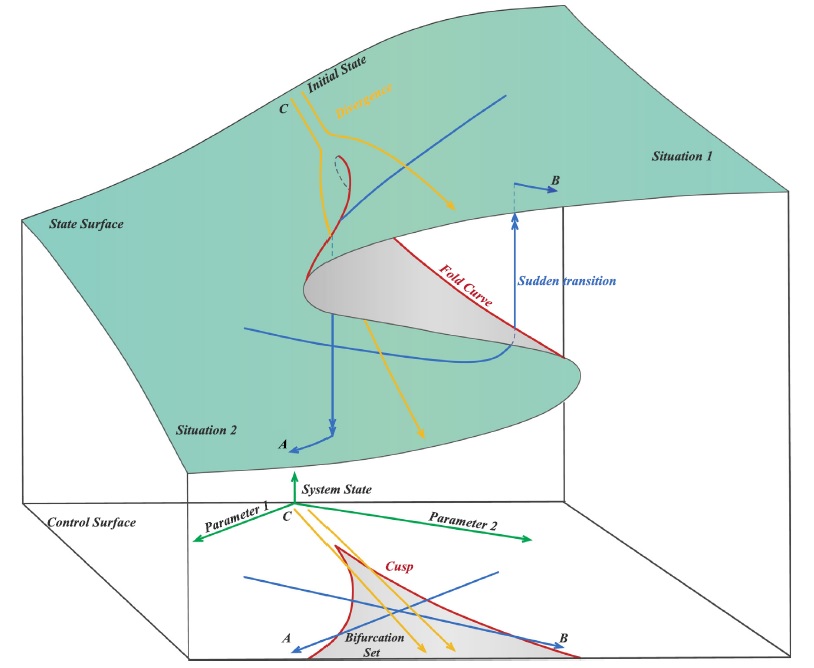
Катастрофа сборки может быть представлена в виде структуры критических точек семейства функций, которая зависит от двух параметров. В катастрофе сборки есть как траектории без перескока, с плавным развитием, так и со скачком в развитии. Структура катастрофы сборки схематично показана на рис. 2. На этом рисунке надпись «State surface» обозначает «поверхность состояния», то есть множество точек состояния системы в параметрическом пространстве управляющих параметров. Надписи «Situation 1» и «Situation 2» – ситуации 1 и 2 – относятся к областям параметрического пространства, в которых может иметь место только одна устойчивая точка равновесия. Проецирование поверхности состояния осуществляется на плоскость «Control surface». Границы бифуркационного множества «Bifurcation set» и границы катастрофы сборки «Fold curve» на рис. 2 показаны красной линией. Серым цветом на плоскости «control surface» показано бифуркационное множество «Bifurcation set». Зелеными стрелками показаны направления изменения управляющих параметров катастрофы сборки. Синими линиями с одинарными стрелками показаны примеры траекторий изменения состояния системы на поверхности состояния и на горизонтальной плоскости «control surface». Синими линиями с двойными стрелками показаны «Sudden transition» – внезапные переходы («перескоки») между состояниями системы. Желтыми линиями со стрелками показаны примеры траекторий изменения состояния системы, демонстрирующие свойство дивергенции – «Divergence». В начальном состоянии – «Initial State» – эти траектории могут располагаться близко друг от друга. Но затем при изменении управляющих параметров они могут существенно расходиться на поверхности состояния и оказываться на разных плоскостях – одна на верхней плоскости, а другая на нижней. Однако проекции этих траекторий на бифуркационное множество остаются такими же близкими, как и в начальном состоянии.
Рис. 2. Структура катастрофы сборки (Wang et al., 2017) (пояснения в тексте) Fig. 2. Structure of cusp catastrophe (Wang et al., 2017) (explanations in the text)
Основополагающим понятием в теории катастроф является понятие «потенциала» системы. Именно значение потенциала (или потенциальной функции) определяет способность изменения текущего состояния системы и возникновение перехода (часто внезапного) в качественно иное состояние. Катастрофические явления претерпевают периоды равновесия, когда потенциал минимален, и периоды внезапных изменений, когда потенциал максимален. Для катастрофы сборки потенциальная функция F (x,a,b) зависит от двух параметров и имеет следующий вид (Постон, Стюарт, 1980; Гилмор, 1984): где x – переменная состояния (в данном исследовании это биомасса фитопланктона), a, b – управляющие параметры (нормальный и расщепляющий соответственно). Параметр a определяет, когда происходит внезапный скачок, а параметр расщепления b регулирует масштаб скачка. Критические вырожденные точки семейства F (x,a,b) находим из условия равенства нулю первой производной функции (1) по переменной состояния x: Бифуркационное множество (bifurcation set на рис. 2) определяет область неустойчивости системы. Пространство управляющих параметров разделяет плоскость (a, b) на несколько областей. Область неустойчивости/устойчивости зависит от знака дискриминанта Для расчета значений управляющих параметров a, b был использован многомерный анализ данных на основе метода главных компонент. Процедуры статистического анализа данных мониторинга выполняли в пакете программ Statistica, v.7. Для периода 2002–2010 гг. суммы значений факторных нагрузок главных компонент рассчитывали для каждой станции мониторинга и каждого года наблюдения. Значения параметров a, b вычисляли через суммы значений факторных нагрузок первых четырех главных компонент. Для параметра a главные компоненты рассчитывали на основе данных по температуре (Tw), солености (Sw) и прозрачности (Sd) воды, pH, БПК5 (BOD5), концентрации растворенного в воде кислорода (O2), проценту насыщения воды кислородом (O2, %) (первая группа данных); для параметра b – на основе данных по содержанию в воде аммонийного азота (N – NH4), азота нитритов (N – NO2) и нитратов (N – NO3), растворенного неорганического (DIN), органического (DON) и общего (Ntot) азота, растворенного неорганического (DIP), органического (DOP) и общего (Ptot) фосфора, соотношениям DIN/DIP, DON/DOP, Ntot /Ptot (вторая группа данных). Суммы значений факторных нагрузок главных компонент для соответствующих переменных будем обозначать следующим образом: TFLПеременная (например, для температуры, солености и прозрачности воды – TFLTw, TFLSw, TFLSd и т.д. для всех других переменных). Для того чтобы устранить влияние размерности, перед выполнением анализа данных методом главных компонент и последующим за ним другим вычислительным процедурам, все переменные были предварительно нормированы (нормализованы). Нормализацию проводили с помощью следующего соотношения: в котором x'i,j – нормализованное значение переменной j в момент времени i; xi,j – фактическое значение переменной j в момент времени i; xj,min и xj,max – соответственно минимальное и максимальное значения переменной j в течение всего рассматриваемого промежутка времени. Запишем (2) в следующем виде: В уравнении (4) x' – нормализованное значение переменной состояния. В качестве переменной состояния рассматривали значения суммарной биомассы фитопланктона. Сделаем замену переменных. Пусть y=x'3, a = k1a', b = k2b' - k3. Тогда (4) можно переписать в следующем виде: В (5) a' и b' – нормализованные значения нормального и расщепляющего параметров, k1, k2, k3 – эмпирические коэффициенты. Замена переменных позволила перейти к линейному уравнению относительно коэффициентов k1, k2, k3 . Уравнение (5) можно переписать в следующей матричной форме: n – число имеющихся наблюдений в ряду данных. С учетом сделанной замены переменных, расчет дискриминанта После того как для каждого года рассматриваемого периода 2002–2010 гг., а также каждой станции мониторинговых наблюдений будут рассчитаны суммы значений факторных нагрузок главных компонент и будет выполнена нормализация всех переменных, можно рассчитать значения a'i и b'i – нормализованные значения нормального и расщепляющего параметров. Для каждого конкретного года расчет проводится по каждому i-му месяцу, в котором одновременно проводилась гидрохимическая и гидробиологическая съемки. Соответствующие расчетные формулы имеют следующий вид: Формулы (8), (9) позволяют также для каждого месяца соответствующего года из рассматриваемого периода 2002–2010 гг. оценивать знак и относительную роль (в %) каждого слагаемого («фактора») с учетом значений и знаков весовых коэффициентов. После того как вычислены соответствующие нормализованные значения a'i и b'i нормального и расщепляющего параметров, можно приступить к следующему этапу – расчету значений управляющих параметров a = k1a' и b = k2b' - k3. Для этого в результате решения матричного уравнения (6) предварительно нужно рассчитать значения эмпирических коэффициентов k1, k2, k3. Вектор неизвестных значений коэффициентов k1, k2, k3 в (6) можно найти, например, с помощью метода наименьших квадратов.
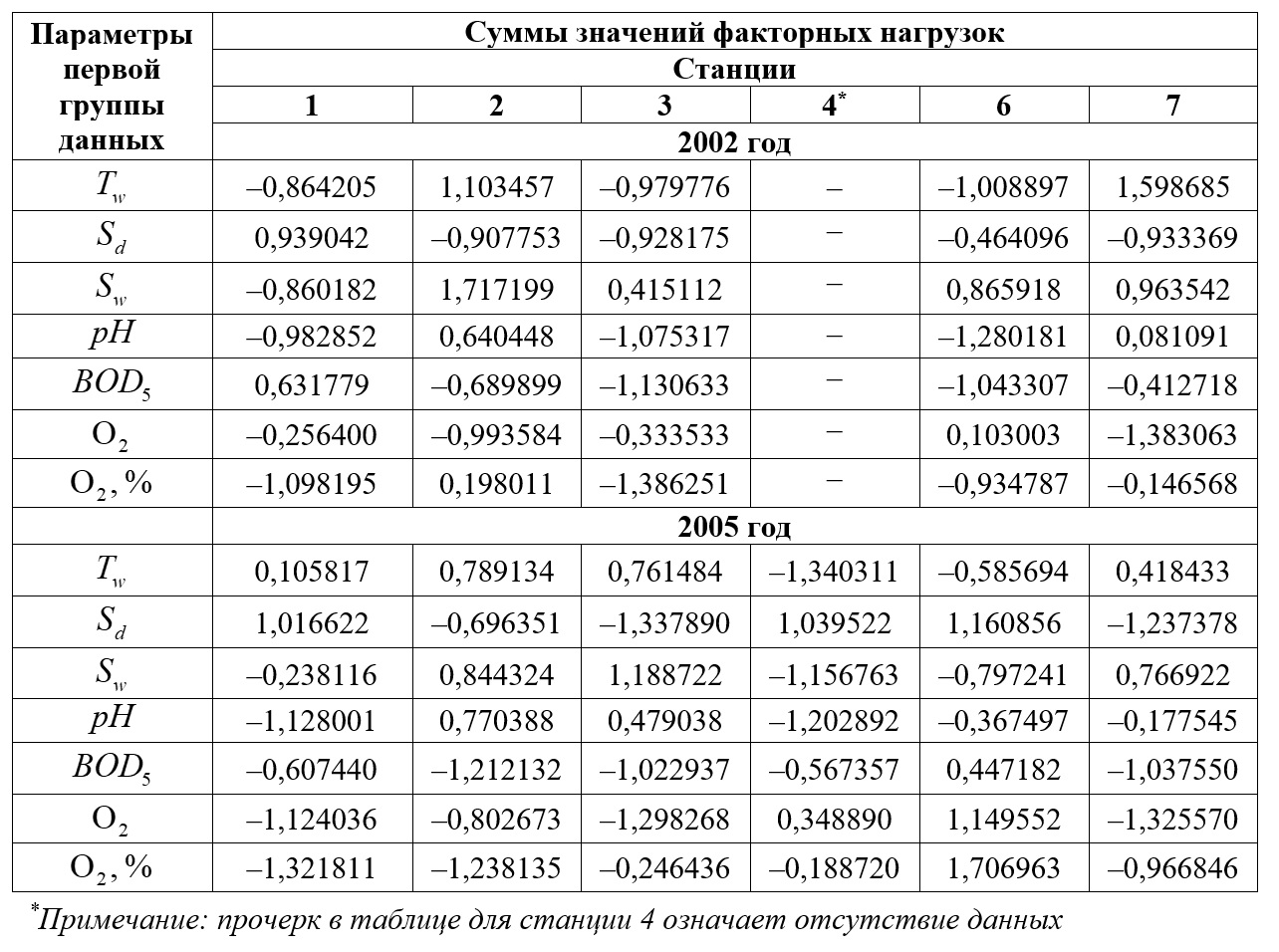
Результаты Анализ данных регулярного ежемесячного мониторинга показал, что для первой группы данных в течение 2002–2010 гг. первые четыре компоненты берут на себя свыше 96% общей дисперсии. Для второй группы данных четырьмя главными компонентами можно охватить свыше 93% общей дисперсии. В результате проведенного исследования было установлено, что знаки суммы значений факторных нагрузок главных компонент для обеих групп данных (параметров гидрологического и гидрохимического режима) могут быть как положительными, так и отрицательными. Значения суммарных факторных нагрузок по абсолютной величине меняются в достаточно широких пределах. В качестве примера в табл. 1 и 2 приведены соответствующие данные для 2002 и 2005 гг. Выявить какие-то устойчивые закономерности их изменений не удается – ни в зависимости от станции, ни в зависимости от года проведения наблюдений. В разные годы на разных станциях наиболее значимыми могут быть свои группы факторных нагрузок, а их вклады в значения соответствующих бифуркационных параметров могут иметь разные знаки. Это позволяет говорить о том, что в многомерном факторном пространстве имеет место существование разнонаправленных градиентов. Воздействия знакопеременных факторов определяют динамику биотической компоненты экосистемы (в данном случае – биомассы фитопланктона). Высокие (по модулю) значения определенных факторных нагрузок по тому или иному параметру свидетельствуют о высоком вкладе данного параметра в многомерный градиент.
Таблица 2. Примеры расчета сумм значений факторных нагрузок первых четырех главных компонент для второй группы данных многолетних наблюдений в 2002 и 2005 гг.
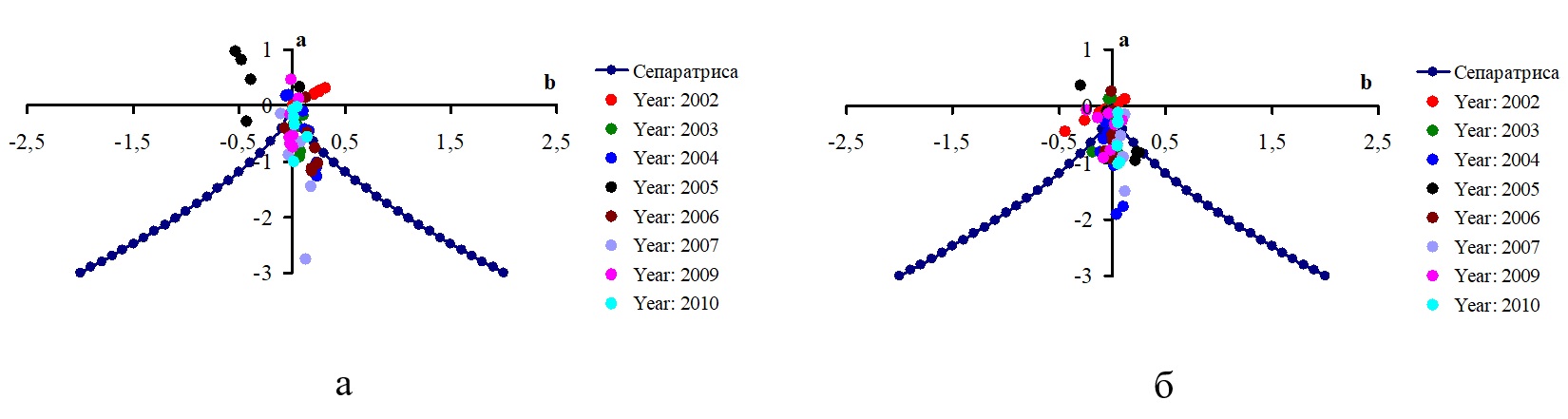
Не существует какого-то единственного сценария «потери устойчивости» в системе, появления бифуркции, последующего скачкообразного перехода из одного состояния устойчивого равновесия в другое. Также нельзя заранее однозначно ответить на вопрос о том, какие конкретно факторы будут влиять на изменения значений управляющих параметров a, b, которые обусловят обратный выход системы за пределы бифуркационного множества и приведут только лишь к последующим плавным изменениям переменной состояния x. Особое внимание следует обратить на такие значения суммарных факторных нагрузок, которые либо выходят за пределы доверительных интервалов, либо являются «выбросами» в статистических распределениях. Именно такие, экстремальные значения, при условии, что совокупность других нагрузок не сможет «сбалансировать» влияние этого «фактора», или же совокупное влияние достаточно значительных по величине «факторов» одного знака могут приводить к кардинальному изменению состояния и последующей динамики переменной состояния системы (т.е. значения суммарной биомассы фитопланктона). В одних случаях это может стать причиной резкого, скачкообразного изменения состояния системы, а в других – обеспечить обратный выход системы за пределы бифуркационного множества и последующие за этим плавные изменения переменной состояния. После того, как вычислены значения параметров a, b, становится возможным рассмотреть вопрос о том, как может меняться положение проекций точек фазового пространства на плоскость (a, b) управляющих параметров катастрофы сборки для различных станций Калининградского (Вислинского) залива Балтийского моря за период 2002–2010 гг. Примеры таких проекций показаны на рис. 3.
Fig. 3. Examples of phase space point projections onto the plane (a, b) of cusp catastrophe control parameters and their position relative to the bifurcation set for different stations of the Kaliningrad (Vistula) Lagoon of the Baltic Sea for the period 2002–2010: а – station 1; б – station 7
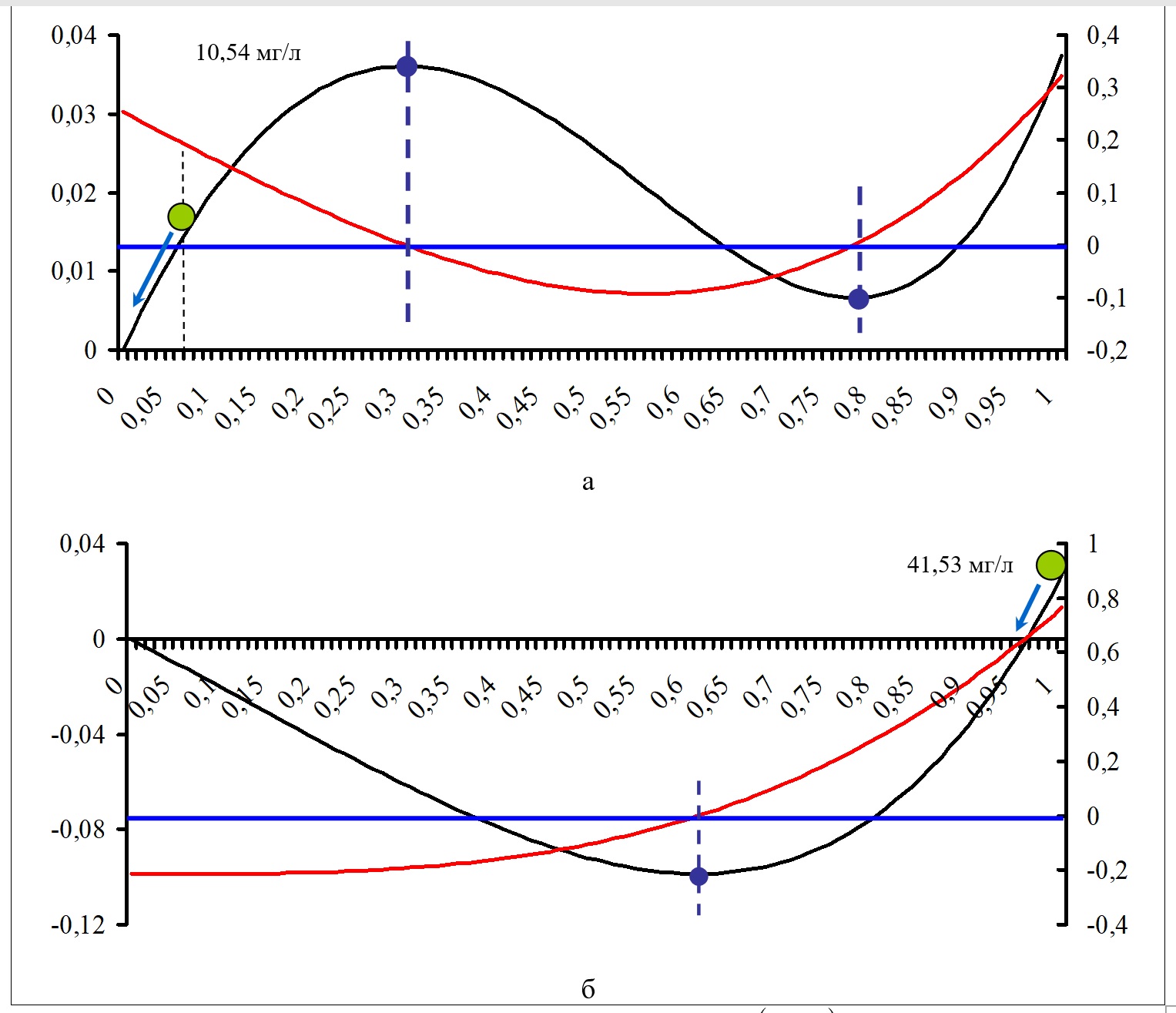
На рис. 4 показаны примеры наиболее типичного вида потенциальной функции F (x,a,b) и ее первой производной для неустойчивого (а) и устойчивого (б) состояния. Расчеты выполняли для конкретных значений управляющих параметров в зависимости от изменения нормализованных значений биомассы фитопланктона (отложены по горизонтальной оси). Потенциальная функция показана линией черного цвета (ее значения отложены на левой вертикальной оси), ее производная – красной линией (ее значения отложены на правой вертикальной оси). Положения максимума и минимума соответствуют нулевым значениям производной и показаны темными кружками. Значения нормализованной биомассы фитопланктона показаны кружком зеленого цвета, а для фактической биомассы приведены ее значения в мг/л. Стрелкой показано направление изменения текущего положения переменной состояния.
Рис. 4. Примеры типичного вида потенциальной функции F (x,a,b) и ее первой производной для неустойчивого (а) и устойчивого (б) состояния (пояснения в тексте) Fig. 4. Examples of the form of a potential function F (x,a,b) and its first derivative for an unstable (a) and stable (b) state (explanations in the text)
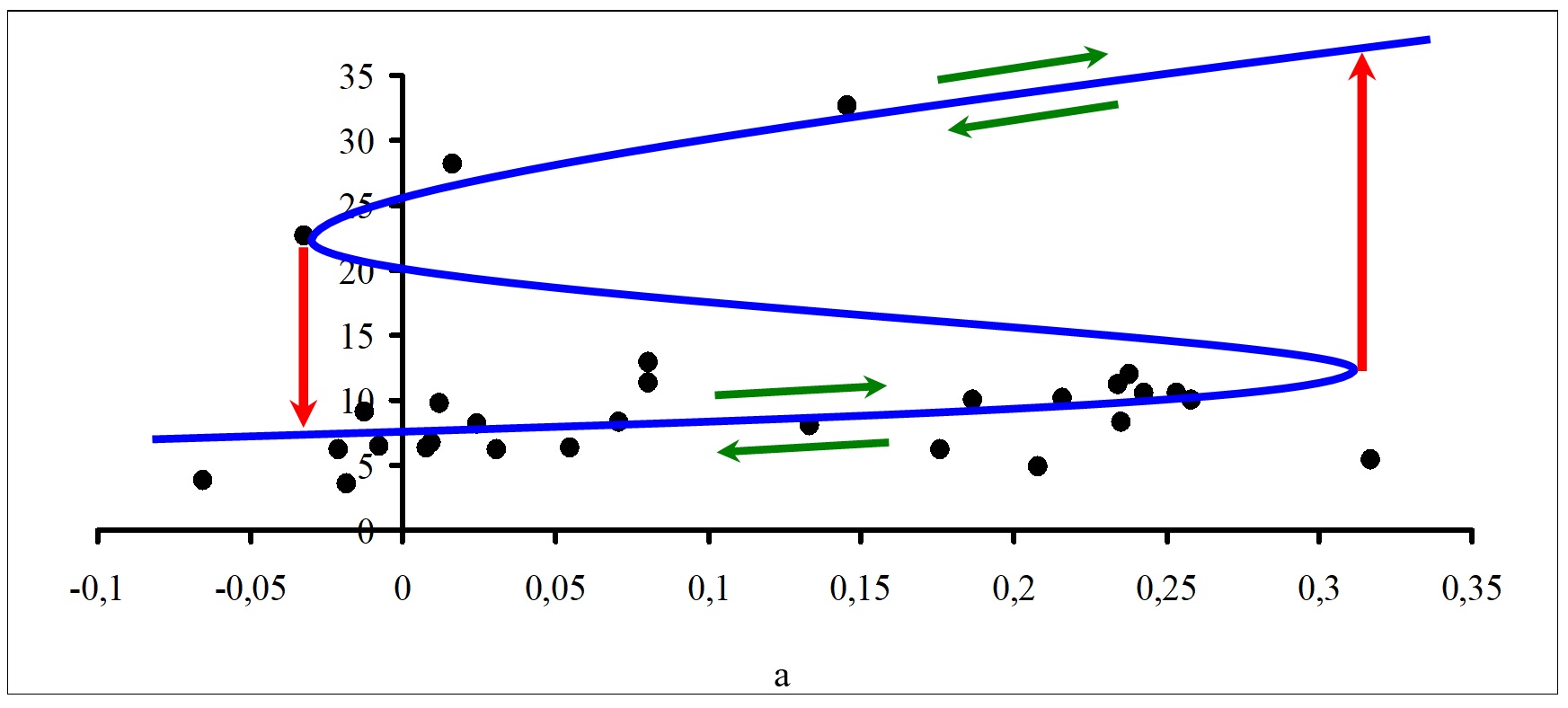
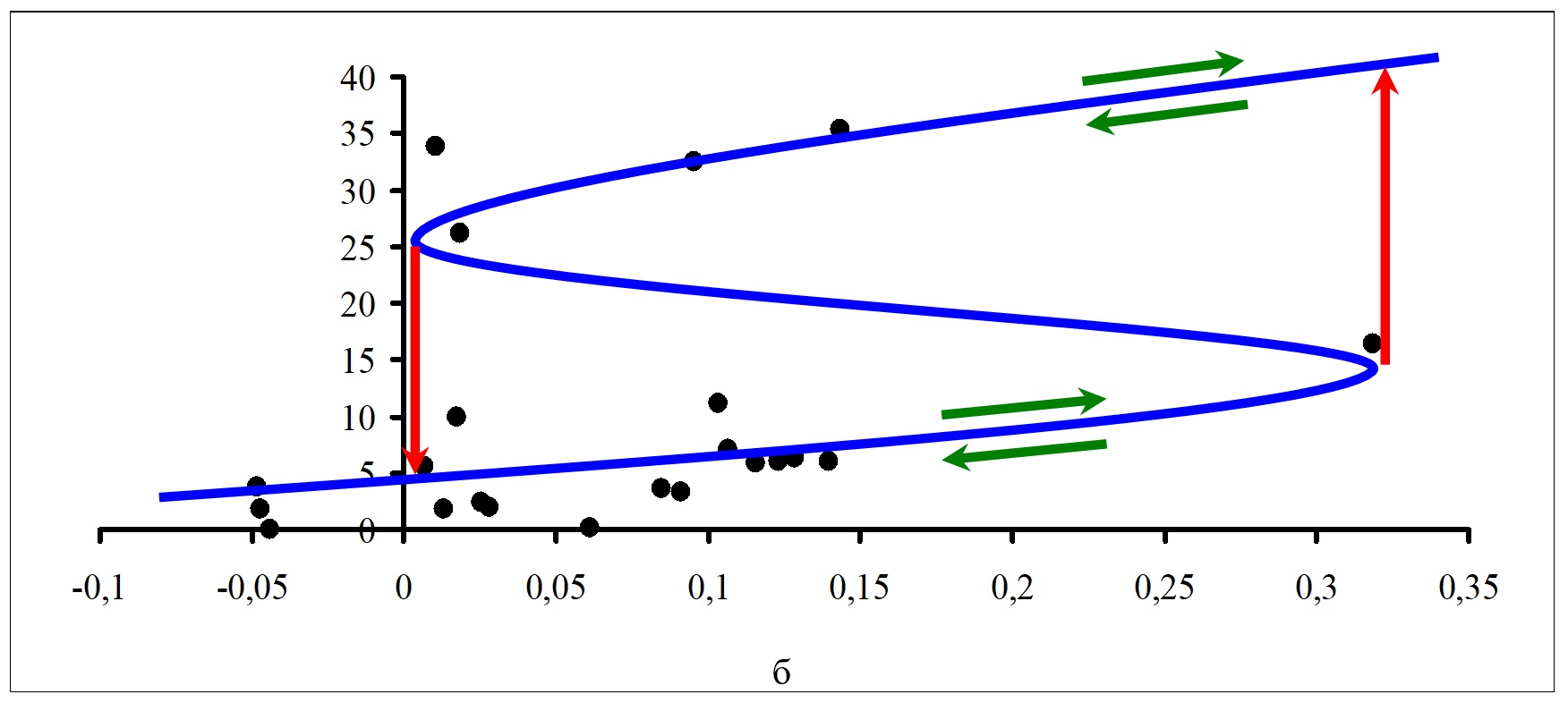
Можно выделить две качественно разные ситуации. Во-первых, существуют станции и отдельные годы, для которых в течение вегетационного сезона все проекции точек фазового пространства на плоскость (a, b) находятся за пределами бифуркационного множества. Например, такая ситуация имела место для станции 1 в 2005 г., для станции 7 в 2002 г. В этих случаях каждой точке (a, b) в пространстве параметров соответствует только одно состояние равновесия, которое всегда устойчиво. Таким образом, изменения значений управляющих параметров a, b способны приводить только лишь к плавным изменениям суммарной биомассы фитопланктона с течением времени под влиянием комбинации факторов окружающей среды. Во-вторых, существуют станции и годы, для которых в течение вегетационного сезона одна часть проекций точек фазового пространства на плоскость (a, b) находится внутри бифуркационного множества, а другая часть – за его пределами. Это свидетельствует о том, что точкам (a, b) в пространстве параметров соответствуют качественно разные состояния равновесия фитопланктонного звена водной экосистемы, которые могут быть как устойчивыми, так и неустойчивыми. Таким образом, изменения значений управляющих параметров a, b в этом случае будут приводить не только к плавным, но также и к резким, скачкообразным изменениям суммарной биомассы фитопланктона с течением времени под влиянием определенной, складывающейся на данный момент времени комбинации факторов окружающей среды. Результатом такого изменения становится не разрушение и/или гибель, а динамический переход системы к качественно иной траектории ее развития. Предсказание итога такого перехода является трудноразрешимой задачей. Рассмотрим примеры многолетнего изменения биомассы фитопланктона российской части Вислинского залива Балтийского моря с точки зрения модели катастрофы сборки. Примеры структуры катастрофы сборки для биомассы фитопланктона для станций 1 и 6 за период 2002–2010 гг. в зависимости от изменения значений управляющего параметра расщепления b показаны на рис. 5. Аналогичные графики были получены и для всех других станций проведения мониторинга. Значения биомассы фитопланктона (мг/л) отложены по вертикальной оси. Значения управляющего параметра b (в безразмерных единицах) отложены по горизонтальной оси. Синей линией показано положение состояний равновесия, черными кружками – значения суммарной биомассы при разных значениях параметра расщепления . Зелеными стрелками показаны возможные направления изменения состояния биотической компоненты экосистемы вдоль состояний равновесия, а красными стрелками – направления возможных «перескоков» из одного устойчивого состояния в другое. Для построения графиков рис. 5 для каждой станции проведения наблюдений выбирали те значения биомассы, которые соответствовали отрицательным значениям дискриминанта. При более частом проведении гидробиологических съемок аналогичные графики можно построить отдельно для каждого года. Как следует из формулы (9), при расчете значений расщепляющего параметра b используются данные по содержанию форм биогенных элементов в водной среде и соотношений между ними. Таким образом, именно эти факторы (по отдельности или в совокупности) в определенных случаях могут приводить к резкому, скачкообразному изменению значений суммарной биомассы фитопланктона (как в сторону ее увеличения, так и в сторону уменьшения), а в других – обеспечивают изменения состояния биотической компоненты экосистемы только вдоль состояний равновесия. Заметим, что амплитуда скачка будет зависеть также и от текущих значений нормального управляющего параметра a.
Рис. 5. Примеры структуры катастрофы сборки для биомассы фитопланктона за период 2002–2010 гг. в зависимости от изменения значений параметра расщепления b для дискриминанта D<0: Fig. 5. Examples of the cusp catastrophe structure for phytoplankton biomass for the period 2002–2010 depending on the change in the values of the splitting parameter b for discriminant D<0:
Исследование положений равновесия само по себе не может сказать нам однозначно, где и при каких именно комбинациях факторов будет происходить «прыжок», т.е. скачкообразное изменение переменной состояния. Вислинский залив Балтийского моря, несмотря на свои небольшие размеры, отличается существенной пространственной неоднородностью гидрологических, гидрохимических и гидробиологических параметров. Наиболее заметно неоднородность проявляется в весенний период, а также при затоках морских вод в залив и стоке воды из залива в море. Поэтому для каждой станции мониторинга в конкретный момент времени могут быть свои ключевые комбинации факторов, которые влияют на текущее состояние устойчивого/неустойчивого равновесия.
Заключение Полученные в данной работе результаты показывают, что фитопланктонное звено экосистемы Вислинского залива может иметь несколько состояний равновесия – как устойчивых, так и неустойчивых. Соответственно этому, обязательно должны существовать критические точки (critical tipping points), в которых может происходить резкая смена динамических режимов функционирования экосистемы. Это может приводить к тому, что в зависимости от изменения текущих значений управляющих параметров экосистема Вислинского залива потенциально может находиться в разных своих состояниях, а также демонстрировать сложную временную динамику и внезапно «перескакивать» между устойчивыми состояниями. Структурно-функциональные сдвиги в экосистеме могут быть плавными, постепенными, а резкие изменения структуры системы (если они произошли) обратимыми. Обратимость структурных изменений экосистемы и характера состояний динамического равновесия не гарантирует того, что временные изменения переменных состояния системы (в данной работе это суммарная биомасса фитопланктона) также будут обратимыми. Это означает, что при изменении значений управляющих параметров на точные противоположные их значения система не обязательно вновь перейдет в первоначальное свое состояние. Одни и те же значения управляющих параметров a, b, а также значения дискриминанта D=4a3 + 27b2 могут быть получены совершенно разными комбинациями факторных нагрузок, которые с течением времени меняют свой вес и/или знак. Знаки и значения дискриминанта можно использовать для анализа, оценки текущего состояния и построения статистического прогноза устойчивости/неустойчивости системы или отдельных ее частей под влиянием изменений факторов окружающей среды. Предложенная в данной работе методика использования теории катастроф для анализа сезонной и межгодовой динамики фитопланктона в Вислинском заливе представляет собой только первый этап работы. Дальнейшие исследования обязательно будут учитывать видовой состав с выделением наиболее важных функциональных групп фитопланктона. Это позволит более детально исследовать комбинации «ведущих» факторов, а также учесть сезонные закономерности, связанные с доминированием разных систематических или функциональных групп фитопланктона в те или иные периоды их вегетационной активности.
Исследования выполнены в рамках государственного задания ФГБУ «ВНИРО» Атлантического филиала ВНИРО («АтлантНИРО») № 076-00007-22-00 часть II, раздел 5 и в рамках государственного задания Института океанологии им. П.П. Ширшова РАН № FMWE-2021-0007. Авторы заявляют об отсутствии конфликта интересов, требующего раскрытия в данной статье. Благодарности. Авторы считают своим приятным долгом выразить слова благодарности и глубокой признательности всем анонимным рецензентам, которые взяли на себя нелегкий труд прочтения и критического анализа статьи. Высказанные ими замечания и предложения были важны для нас и по возможности учтены.
Список литературы
Статья поступила в редакцию 17.06.2022
Об авторах Подгорный Константин Алексеевич – Konstantin A. Podgornyy кандидат физико-математических наук kapborok@mail.ru Дмитриева Ольга Александровна – Olga A. Dmitrieva кандидат биологических наук phytob@yandex.ru Корреспондентский адрес: 236022, Калининград, ул. Дмитрия Донского, 3, АтлантНИРО; тел. +7(4012)21-99-97.
ССЫЛКА: Подгорный К.А., Дмитриева О.А. Опыт использования методов теории катастроф при описании динамики фитопланктона в Вислинском заливе Балтийского моря // Вопросы современной альгологии. 2022. № 2 (29). С. 1–14. URL: http://algology.ru/1822 DOI – https://doi.org/10.33624/2311-0147-2022-2(29)-1-14 EDN – AHNBND
Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно.
Experience in using the methods of catastrophe theory in describing the dynamics Konstantin A. Podgornyy1, Olga A. Dmitrieva1,2 1Atlantic Branch of VNIRO – ”AtlantNIRO” (Kaliningrad, Russia) Ideas and methods of catastrophe theory are widely used to analyze “regime shifts” in ecosystems. In this work, the analysis of the seasonal dynamics of phytoplankton in the Russian part of the Vistula Lagoon of the Baltic Sea is carried out on the basis of using the simplest possible model of the catastrophe theory − cusp catastrophe. The total phytoplankton biomass is considered as a state variable. The use of one of the methods of multivariate analysis − principal components method − made it possible to carry out the entire analysis depending on a small number of parameters. A calculation of the control parameters values is carried out using the factor loadings of the main components for the data that characterize the hydro-physical and hydro-chemical regimes as well as the level of biogenic loading on the aquatic ecosystem of the lagoon. It was found that, depending on the specific situation, the same factor or different combinations of factors can play both a stabilizing role and be the cause of destabilization. Key words: aquatic ecosystems; integrated analysis; mathematical modeling; regime shift; catastrophe theory; cusp catastrophe; sustainability of ecosystems
References
Authors Podgornyy Konstantin A. ORCID – https://orcid.org/0000-0001-7402-6324 Atlantic Branch of VNIRO (”AtlantNIRO”), Kaliningrad, Russia kapborok@mail.ru Dmitrieva Olga A. ORCID – https://orcid.org/0000-0002-4808-3728 Atlantic Branch of VNIRO (”AtlantNIRO”), Kaliningrad, Russia; phytob@yandex.ru
ARTICLE LINK: Podgornyy K.A., Dmitrieva O.A. Experience in using the methods of catastrophe theory in describing the dynamics of phytoplankton in the Vistula lagoon of the Baltic Sea. Voprosy sovremennoi algologii (Issues of modern algology). 2022. № 2 (29). P. 1–14. URL: http://algology.ru/1822 DOI – https://doi.org/10.33624/2311-0147-2022-2(29)-1-14 EDN – AHNBND When reprinting a link to the site is required
На ГЛАВНУЮ

|
|||
|
| ||