|
|
|
|
с международным участием «Водоросли: проблемы таксономии и экологии, использование в мониторинге и биотехнологии» (12–18 сентября 2022, Москва, Россия) Пространственная изменчивость видового и функционального состава водорослей перифитона в озерах Карельского перешейка
Русанов А.Г., Станиславская Е.В. Alexander G. Rusanov, Elena V. Stanislavskaya
Санкт-Петербургский ФИЦ РАН, Институт озероведения РАН
УДК 574.586
Изменчивость видовой и функциональной структуры водорослей перифитона в зависимости от гидрохимических, физических, климатических и пространственных факторов исследована в 55 озерах Карельского перешейка. Для оценки факторов, влияющих на функциональную структуру, были выделены восемь функциональных групп, различающихся морфологией клетки/колонии и характером прикрепления к субстрату. Анализ компонент дисперсии показал, что факторы среды (гидрохимические и климатические) преобладали над пространственными переменными в объяснении изменчивости видовой структуры. Напротив, функциональная структура больше зависела от пространственных переменных, чем от факторов среды (гидрохимических и физических). Это говорит о том, что процессы расселения, зависящие от особенностей ландшафта региона и гидрологической связности отдельных речных сетей, играют более заметную роль в пространственной изменчивости функционального состава перифитона. Ключевые слова: перифитон; функциональные группы; теория метасообщества; расселение организмов; факторы среды; экологический мониторинг
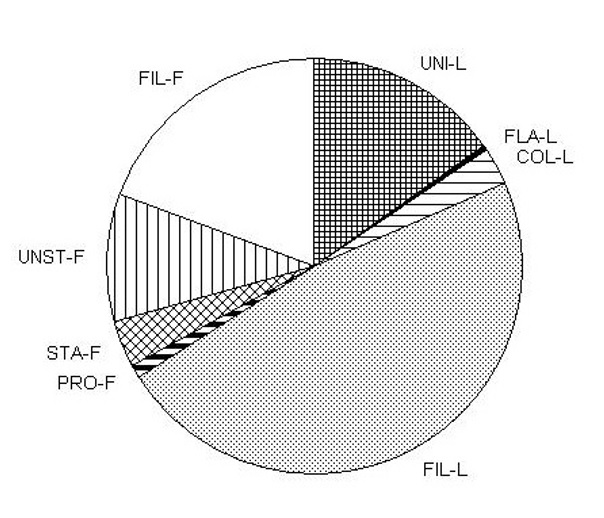
В последние десятилетия в экологии сообществ возобладал подход, рассматривающий пространственно-временное распределение организмов с позиции теории метасообщества, согласно которой формирование видовой структуры обусловлено совместным влиянием локальных факторов среды, связанных с дифференциацией ниш, и процессов расселения видов (Leibold et al., 2004). Метасообщество определяется как совокупность локальных видовых группировок (сообществ), которые связаны друг с другом посредством расселения множества потенциально взаимодействующих видов (Wilson, 1992). Относительная роль факторов среды и процессов расселения в регуляции структуры сообщества, главным образом зависит от пространственной протяженности изучаемой территории (Heino, 2013). Согласно теории метасообщества, влияние факторов среды на структуру сообщества максимально при исследованиях среднего масштаба (102–105 км2), когда скорость расселения не слишком низкая и не слишком высокая, и большинство организмов могут достичь местообитаний с оптимальными условиями среды. Роль ограниченного расселения возрастает при увеличении масштаба исследования (от 105 км2 до континентального), при котором значительные расстояния, географические барьеры и низкая способность к расселению мешает видам эффективно реагировать на изменение условий среды. Наоборот, роль избыточного расселения преобладает в исследованиях малого масштаба (1–102 км2), когда непрерывная миграция организмов из соседних местообитаний приводит к существованию видов в условиях, далеких от их видоспецифического оптимума. Эмпирические исследования, проведенные в разнотипных водных экосистемах, подтверждают влияние масштаба исследования на относительную важность факторов среды и расселения в формировании структуры водорослевых сообществ (Soininen et al., 2011; Tang et al., 2013; Soininen et al., 2016; Rusanov et al., 2022). В то же время, следует учитывать, что в исследованиях среднего масштаба сообщества могут испытывать одновременное влияние ограниченного и избыточного расселения, значимость которых меняется в зависимости от ландшафтной конфигурации региона, связности гидрографической сети и расселительной способности отдельных групп видов (Göthe et al., 2013; Dong et al., 2016; Lansac-Tôha et al., 2016). В настоящей работе оценивалось влияние факторов среды (гидрохимических, физических и климатических) и процессов расселения на изменчивость видовой и функциональной структуры водорослей перифитона в озерах Карельского перешейка. Целью работы было сравнить относительную важность средовых факторов и процессов расселения в регуляции состава видов и функциональных групп перифитона и определить потенциальную возможность использования функциональных групп в экологическом мониторинге. Отбор проб водорослей перифитона проводили в июле 2009–2019 гг. в 55 озерах Карельского перешейка. Пробы перифитона отбирали с погруженных частей воздушно-водных макрофитов (в основном Phragmites australis). Одновременно отбирали гидрохимические пробы для определения рН, О2 (мг/л), % О2, цветности (Color, град.), БПК (BOD, мг О2/л), ХПК (COD, мг О2/л), общего азота (TN, мг/л), общего фосфора (TP, мкг/л) и N/P. Из физических факторов учитывались площадь водоема (S, км2), средняя глубина (Hm, м), максимальная глубина (Hmах, м), глубина Секки (Secchi, м) и динамическое отношение (S/Hm) (Hakanson, 1982). Климатические переменные были взяты с сервиса WorldClim (https://www.worldclim.org/) (Harris et al., 2014), на котором представлены климатические данные за период 1901–2019 гг. на координатной сетке высокого пространственного разрешения (0.5° шир./долг.). Учитывались среднегодовая температура воздуха (T.my, °C), средняя температура за летний сезон (T.ms, °C), температура в июле (T.j, °C), годовая изменчивость температуры (T.SD), среднегодовой уровень осадков (P.my, мм), средний уровень осадков за летний сезон (P.ms, мм), уровень осадков в июле (P.j, мм) и годовой коэффициент вариации осадков (P.CV). Пространственные переменные, являющиеся прокси-переменными для оценки процессов расселения, были сгенерированы при помощи PCNM-метода (principle coordinates of neigbour matrices, Dray et al., 2006). Путем усечения матрицы Эвклидовых расстояний между географическими координатами озер было получено 33 пространственных переменных, позволяющих учесть широкий диапазон пространственных зависимостей. Первые широкомасштабные переменные отражают зависимости в масштабе всего района исследования, моделируя влияние ограниченного расселения. Напротив, последние узко-масштабные переменные, отражающие зависимости между соседними близко расположенными озерами, используются для моделирования эффекта избыточного расселения (Heino et al., 2015). В пробах перифитона было определено 262 вида водорослей. Наибольшее число видов составляли Chlorophyta (116 видов), за которыми следовали Bacillariophyta (92 вида), Cyanoprokaryota (39 видов), Euglenophyta (10 видов), Xanthophyta (3 вида) и Dinophyta (2 вида). Водоросли перифитона обладают широким разнообразием жизненных форм, определяющих их адаптивные стратегии и специфическое расположение в слое перифитона (Biggs et al., 1998). Согласно эколого-морфологическим признакам, все водоросли были разделены на две группы с 4-мя подгруппами (всего 8 функциональных групп): слабо прикрепленные (одноклеточные (UNI-L), жгутиковые (FLA-L), колониальные (COL-L) и нитчатые (FIL-L)) и прочно прикрепленные (распростертые (PRO-F), стебельчатые (STA-F), безстебельчатые (UNST-F) и нитчатые (FIL-F)). Данное разделение на функциональные группы основано на признаках, которые могут быть отнесены к трем основным функциональным «размерностям», связанным со 1) способностью к движению, 2) способностью переносить неблагоприятные условия окружающей среды и 3) способностью эффективно использовать ресурсы в конкуренции с другими видами (Bauer et al., 2021). По числу видов лидировали UNI-L (105 видов), за которыми следовали FIL-L (53), FIL-F (31), COL-L (24), STA-F (21), UNST-F (13), FLA-L (12) и PRO-F (6). Функциональные группы также различались средним относительным обилием (рис. 1). Наибольшим относительным обилием (% от общей численности) отличались FIL-L (47,6%); за ними в порядке убывания следовали FIL-F (19,4%), UNI-L (15,2%), UNST-F (10%), STA-F (3,6%), COL-L (2,9%), PRO-F (1%) и FLA-L (0,4%). Наиболее обильными видами в группах были: Epithemia adnata (Kützing) Brébisson (5,0%, UNI-L); Trachelomonas hispida (Perty) F.Stein (0,1%, FLA-L) и T. volvocina (Ehrenberg) Ehrenberg (0,1%, FLA-L); Merismopedia tenuissima Lemmermann (1,0%, COL-L); Tabellaria flocculosa (Roth) Kützing (12,5%, FIL-L) и T. fenestrata (Lyngbye) Kützing (11,1%, FIL-L); Cocconeis pediculus Ehrenberg (0,4%, PRO-F); Gomphonema truncatum Ehrenberg (0,7%, STA-F) и Cymbella cistula (Ehrenberg) O.Kirchner (0,6%, STA-F); Fragilaria capucina Desmazières (6,7%, UNST-F); Oedogonium spp. (9,2%, FIL-F). Рис. 1. Среднее относительное обилие функциональных групп перифитона
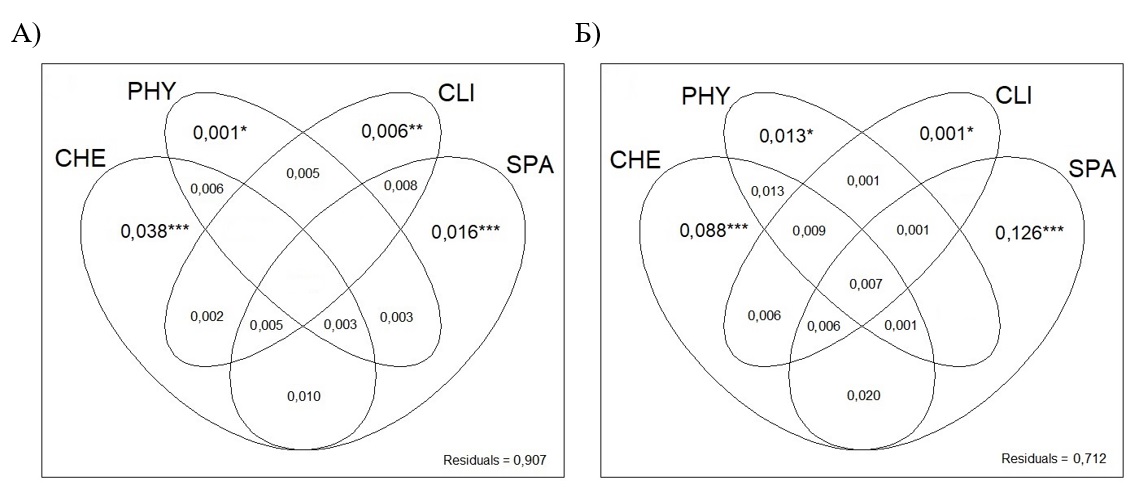
Чтобы оценить отклик видовой и функциональной структуры водорослей перифитона на гидрохимические, физические, климатические и пространственные переменные и определить фракции в изменчивости биологических данных, приходящиеся на каждый набор предикторов и их ковариацию, проводили анализ компонент дисперсии на основе парциального анализа избыточности (partial redundancy analysis, Borcard et al., 1992). Первоначальный отбор переменных, оказавших статистически значимый (p ≤ 0,05) эффект на структуру перифитона, проводили при помощи процедуры прямой селекции (forward selection, Blanchet et al., 2008). Оценивали коэффициенты детерминации R2 моделей для каждого набора предикторов; статистическая значимость их чистых фракций в объясненной изменчивости без учета взаимодействия между ними тестировалась при p ≤ 0,05. Видовая и функциональная структура сообщества была выражена как относительная численность видов и функциональных групп, соответственно. Анализ данных проводили при помощи статистического пакета vegan (Oksanen et al., 2019) в среде R ver. 3.6.3 (https://cran.rproject.org/). Анализ компонент дисперсии для видового состава перифитона показал, что гидрохимические (рН, цветность, % О2, ХПК) и климатические (T.j, T.SD) факторы вместе отвечают за основную долю (5%) вариабельности данных, тогда как вклад физических факторов (S) незначителен (0,1%) (рис. 2А, табл. 1). Набор пространственных предикторов в основном включает широкомасштабные переменные (PCNM 3, 4, 11), указывая на возможный эффект ограниченного расселения. Однако, поскольку вклад пространственных переменных меньше вклада гидрохимических и климатических факторов, составляя всего 2%, условия среды являются основными драйверами пространственной изменчивости видовой структуры перифитона.
Рис. 2. Результаты анализа компонент дисперсии, показывающие относительный вклад гидрохимических (CHE), физических (PHY), климатических (CLI) и пространственных (SPA) переменных в изменчивость состава видов (А) и функциональных групп (Б). Значения представляют R2-значения, скорректированные на количество предикторов в модели. Отрицательные значения фракций не показаны. Статистическая значимость: * p ≤ 0,05; ** p ≤ 0,01; *** p ≤ 0,001.
Анализ компонент дисперсии для функционального состава перифитона показал, что гидрохимические (TP, О2, % О2, БПК) и физические (S) факторы в совокупности отвечают за 10% вариабельности данных, а вклад климатических факторов (T.SD) незначителен (0,1%) (рис. 2Б, табл. 1). Доля пространственных переменных составляет 13%, превышая суммарную долю факторов среды. При этом вклад широкомасштабных пространственных переменных (PCNM 5, 11) равен вкладу узко-масштабных переменных (PCNM 22, 28). Это говорит о том, что пространственное распределение функциональных групп перифитона в значительной степени зависит как от влияния ограниченного расселения между отдаленными озерами из несвязанных речных бассейнов, так и от влияния избыточного расселения между соседними водоемами, принадлежащими к одному бассейну.
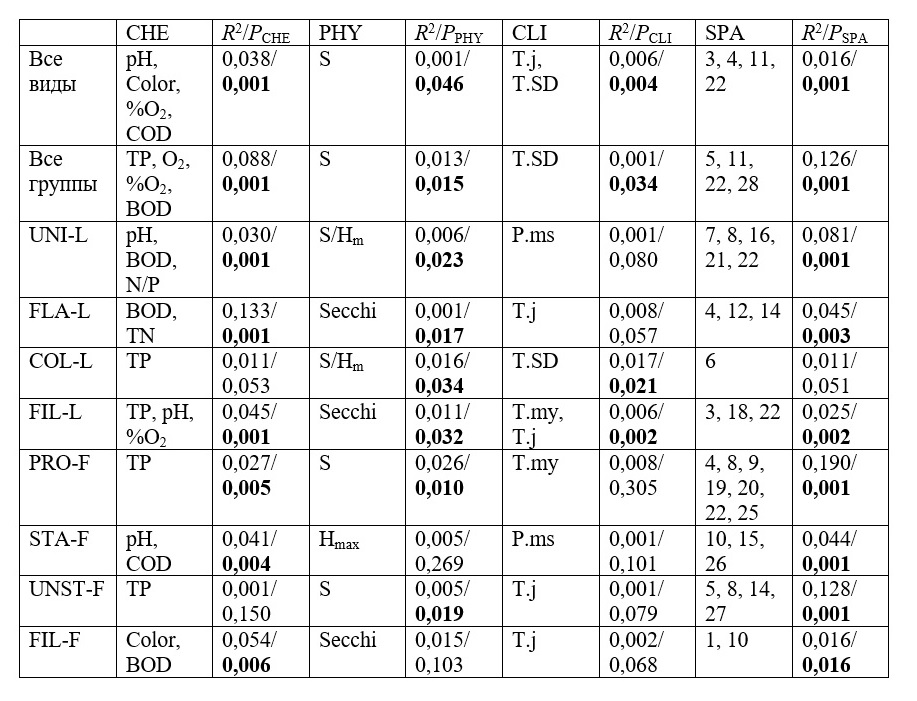
Таблица 1. Результаты парциального анализа избыточности для полного видового состава (все виды), полного функционального состава (все группы) и отдельно для каждой функциональной группы сообщества водорослей перифитона. Все виды перифитона делятся на слабо прикрепленные (одноклеточные (UNI-L), жгутиковые (FLA-L), колониальные (COL-L) и нитчатые (FIL-L)) и прочно прикрепленные (распростертые (PRO-F), стебельчатые (STA-F), безстебельчатые (UNST-F) и нитчатые (FIL-F)). Table 1. Results of the partial redundancy analysis for the full species composition (all taxa), the full functional composition (all groups), and separately for each functional group of the periphytic algal community. All species were divided into loosely attached (unicellular (UNI-L), flagellate (FLA-L), colonial (COL-L) and filamentous (FIL-L)) and firmly attached (prostrate (PRO-F), with stalk (STA-F), without stalk (UNST-F) and filamentous (FIL-F)).
Примечания. Notes.
Пространственное распределение функциональных групп перифитона также подчиняется закономерностям, связанным с характером прикрепления водорослей к субстрату, и следовательно, с их способностью к расселению. Прочно прикрепленные распростертые (PRO-F) и безстебельчатые (UNST-F) водоросли показали наиболее сильную зависимость от широкомасштабных пространственных переменных (табл. 1), что говорит об эффекте ограниченного расселения и хорошо согласуется с их более слабой способностью к расселению. С другой стороны, слабо прикрепленные одноклеточные водоросли (UNI-L), показавшие сильную зависимость от пространства (табл. 1), характеризуются высоким вкладом узко-масштабных переменных. Это указывает на эффект избыточного расселения, который испытывают слабо прикрепленные водоросли с высокой способностью к расселению. Таким образом, проведенный анализ показал, что связанные с расселением процессы вносят значительный вклад в формирование функциональной структуры сообщества перифитона Карельского перешейка, что необходимо учитывать при использовании функциональных групп в экологическом мониторинге.
Работа выполнена в рамках темы Института озероведения РАН № 0154-2019-0001. Авторы заявляют об отсутствии конфликта интересов, требующего раскрытия в данной статье.
Список литературы
Статья поступила в редакцию 21.06.2022
Об авторах Русанов Александр Геннадьевич – Alexander G. Rusanov кандидат биологических наук a_rusanov@yahoo.com Станиславская Елена Владимировна – Elena V. Stanislavskaya кандидат биологических наук, доцент stanlen@mail.ru Корреспондентский адрес: Россия, 196105, г. Санкт-Петербург, ул. Севастьянова, 9, ИНОЗ РАН. Телефон (812)387-02-60.
ССЫЛКА: Русанов А.Г., Станиславская Е.В. Пространственная изменчивость видового и функционального состава водорослей перифитона в озерах Карельского перешейка // Вопросы современной альгологии. 2022. № 2 (29). С. 38–45. URL: http://algology.ru/1827 DOI – https://doi.org/10.33624/2311-0147-2022-2(29)-38-45 EDN – JVMCWY
Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно.
Spatial variation in taxonomic and functional composition of periphytic algae in lakes of Karelian Isthmus Alexander G. Rusanov, Elena V. Stanislavskaya St. Petersburg Federal Research Center RAS, Institute of Limnology RAS (St Petersburg, Russia) Variation in species and functional structure of periphytic algae community in relation to chemical, physical, climatic and spatial factors was studied in 55 lakes of Karelian Isthmus. To assess factors driving functional structure, all algal species were divided into eight functional groups that differ in cell/colony morphology and attachment mode to the substrate. Variance partitioning analysis showed that environmental factors (chemical and climatic) prevailed over spatial variables in explaining variation in periphyton species structure. On the contrary, periphyton functional structure depended more on spatial variables than on environmental factors (chemical and physical). This suggests that dispersal processes which are dependent on landscape characteristics of the region and hydrological connectivity along individual river networks play a more significant role in spatial variation of functional composition of periphytic algae. Key words: periphyton; functional groups; metacommunity theory; dispersal of organisms; environmental factors; ecological monitoring
References
Authors Rusanov Alexander G. ORCID – http://orcid.org/0000-0003-1597-0731 St. Petersburg Federal Research Center RAS, Institute of Limnology RAS, St Petersburg, Russia a_rusanov@yahoo.com Stanislavskaya Elena V. ORCID – http://orcid.org/0000-0002-2897-7636 St. Petersburg Federal Research Center RAS, Institute of Limnology RAS, St Petersburg, Russia stanlen@mail.ru
ARTICLE LINK: Rusanov A.G., Stanislavskaya E.V. Spatial variation in taxonomic and functional composition of periphytic algae in lakes of Karelian Isthmus. Voprosy sovremennoi algologii (Issues of modern algology). 2022. № 2 (29). С. 38–45. URL: http://algology.ru/1827 DOI – https://doi.org/10.33624/2311-0147-2022-2(29)-38-45 EDN – JVMCWY When reprinting a link to the site is required Dear colleagues! If you want to receive the version of the article in PDF format, write to the editor, please and we send it to you with pleasure for free.
На ГЛАВНУЮ
|
|||
|
| ||