 |
Идеи Д.А. Сабинина и их воплощение.
Минеральный состав макрофитов как важнейший показатель их вклада в биогеохимический обмен Мирового океана
D. A. Sabinin's ideas and their embodiment.
Mineral composition of macrophytes is one of the most important indicator of their contribution to a biogeochemical exchange of the World Ocean.
Камнев А.Н.1, Бунькова О.М.2, Богатырев Л.Г.2, Стуколова И.В.1,
Яковлев А.С.2
Alexander N. Kamnev1, Olga M. Bunkova2, Lev G. Bogatyrev2, Irina V. Stukolova1, Alexander S. Yakovlev2
1Московский государственный университет им.М.В.Ломоносова,
биологический факультет
2Московский государственный университет им.М.В.Ломоносова,
факультет почвоведения
УДК 550.47 + 581.1
В работе рассматриваются вопросы, связанные с понятием «биогеохимия» и её историческим становлением как самостоятельной науки. Показана структура современной биогеохимии. Определены её цели и задачи. Дано понятие – биосфера. Определены её границы. Рассматриваются некоторые исторические параллели развития физиологии растений, почвоведения и биогеохимии. Особое внимание уделяется идеям Д.А. Сабинина, который обосновал важность развития биогеохимических исследований в морской среде, а в качестве удобных объектов для этих исследований предложил использовать водные фототрофные организмы, в частности, морские водоросли-макрофиты. Большое внимание уделяется изменениям химического состава прибрежной части Чёрного моря. Это море, включая прибрежную зону, рассматривается, с одной стороны, как удобная модельная биогеохимическая система, а с другой, как стратегически важный регион не только для России, но для Европы и Азии. В работе показаны особенности изменения диапазона минерального состава и концентрационных возможностей почвы, донных отложений и макрофитов. Приведены примеры изменения минерального состава в водных и прибрежно-водных растениях от истока до устья малой реки.
Ключевые слова: биогеохимия; биосфера; фототрофы; водные и прибрежно-водные растения-макрофиты; донные отложения; почва; водная среда; Чёрное море; минеральный состав макрофитов; минеральный состав донных отложений; минеральный состав почвы; минеральный состав водной среды.
Введение
Среди естественных дисциплин биогеохимии принадлежит особое место. Это обусловлено многовековым извечным интересом исследователей к роли живых организмов на земной поверхности. Этот интерес проявлялся последовательно в различных научных дисциплинах. К пониманию роли живых организмов учёные пришли далеко не сразу. Длительное время многие исследователи весьма скептически оценивали роль живых организмов. Им отводилось определённое, но часто ограниченное место в некоторых природных процессах, тогда как более грандиозным на Земле всегда представлялось влияние тектонических процессов, в частности вулканов. Не случайно со времён Ламарка существовало две группы исследователей, одни из которых назывались вулканистами, другие нептунистами. Сам Ламарк принадлежал к группе нептунистов и придавал большое значение роли живых организмов во всех процессах, происходящих на Земле.
Понадобилось длительное время для того, чтобы учёные, в том числе и геологи, признали роль живых организмов не только в современных процессах, но и в геологическом времени. Исключительно важная роль в становлении биогеохимии принадлежит физиологии растений и почвоведению.
Великие химики – физиологи растений, такие как Антуан Лавуазье, Батист Дюма, Жан Батист Буссенго, Юстус фон Либих ещё 170-200 лет назад открыли круговорот газов, заложили основу современных представлений о геохимии углерода и основу изучения циклической миграции минеральных элементов в системе почва – растение – почва, получившей название биологического круговорота.
В 80-х годах XIX века в России Василий Васильевич Докучаев углубил и конкретизировал представление великих физиологов растений о деятельности живых организмов на примере почвенного покрова суши. Он писал о том, что должна быть создана особая наука, занимающаяся вопросами взаимодействия живой и косной материи, подразумевая при этом почвоведение. Признавая приоритет почвоведения, ещё в начале тридцатых годов прошлого столетия выдающийся геохимик А.Е. Ферсман подчёркивал, что из всех наук именно почвоведение играет важную роль для геохимии, обогащая её методами, идеями и фактами. Подчёркивая непосредственную роль В.В. Докучаева, В.И. Вернадский писал о том, что именно работы Василия Васильевича определили весь ход его мыслей и направление работы его лаборатории. Сегодня влияние живых организмов, в частности, фототрофов признается одним из важнейших факторов в развитии почв.
Роль современной биогеохимии заключается в том, что она заполняет конкретными данными общепризнанный тезис о роли живых организмов, но не только в почвообразовании, но и в целом на планете. Почва, которая по образному выражению А.Е. Ферсмана является плёнкой на поверхности земли, в наиболее яркой форме отражает особенности биогеохимических процессов, происходящих в биосфере.
В настоящей работе была предпринята попытка рассмотреть не только некоторые теоретические положения выдающегося отечественного исследователя, физиолога растений Д.А. Сабинина в области минерального питания растений, но и дальнейшее развитие этих положений. Важно подчеркнуть, что работы Д.А. Сабинина, связанные с минеральным питанием растений, имеют одинаковое значение как для физиологии растений, так и биогеохимических исследований наземных и водных экосистем. Надо помнить, что для различных биогеохимических исследований всех экологических систем большое значение имеют вопросы, связанные с продуктивностью и химическим составом живых организмов, закономерностями их минерального питания и характеристиками биогеохимических циклов. Более того, надо подчеркнуть, что биогеохимики, проводившие исследования в наземных экосистемах, всегда обращали внимание на исследования, которые проводились в водных экосистемах, и наоборот. Это обусловлено в первую очередь тем, что в любом случае, исследуя водные или наземные экосистемы, мы находимся в пределах одной системы – биосферы. Интересно отметить, что основоположники биогеохимии В.И. Вернадский и А.Н. Виноградов в своих работах всегда обращались к биогеохимическим процессам, происходящим в гидросфере. Наша работа посвящена исследованию некоторых биогеохимических характеристик наиболее характерных групп водных и прибрежно-водных фототрофных организмов черноморского побережья.
Современный взгляд на биогеохимию
Биогеохимия – междисциплинарная наука, изучающая роль живых организмов в миграции и перераспределении химических элементов на Земле. Её основоположником является выдающийся естествоиспытатель и мыслитель XX века В.И. Вернадский (28.2 [12.3].1863 – 6.1.1945). Теоретической основой этой науки является учение о живом веществе и биосфере.
Термин биосфера ввёл в 1875 г. австрийский геолог Эдуард Зюсс (1831-1914). Под этим термином он понимал сферу обитания живых организмов.
В представлениях и разработках В.И. Вернадского биосферой является наружная оболочка Земли, охваченная геохимической деятельностью живого вещества. Главным отличием в таком понимании термина биосфера является то, что В.И. Вернадский воспринимал биосферу не только как место обитания живых организмов, но как глобальную систему взаимодействия живого и неживого вещества.
Под живым веществом Вернадский понимал все разнообразие живой материи и различных форм жизни, а под косным – материю твердой, жидкой и газовой фаз. Таким образом, по В.И. Вернадскому «биосфера – это единение живой материи и внешней части Земного шара». Современные исследования показывают, что биосфера охватывает слои литосферы, гидросферы и тропосферы суммарной мощностью около 40 км (Башкин, 2004).
Взаимодействие живой и косной систем происходит в форме массообмена химическими элементами. Как считал В.И. Вернадский, именно эти процессы поддерживают «геохимическую организованность» биосферы. Так как все эти процессы осуществляются в результате жизнедеятельности организмов, то они были названы им биогеохимическими.
Известно, что при всем многообразии живых организмов (в силу термодинамической открытости этих живых систем), общим условием их существования является обмен веществ со средой обитания. В результате своих физиологических потребностей, живые организмы поглощают химические элементы. Так как в большинстве случаев это поглощение осуществляется селективно, то это вызывает дифференциацию элементов в окружающей среде. Очень важным аспектом жизнедеятельности организмов является также их газовый обмен. Газообразные метаболиты, поступая в атмосферу, постепенно меняют её состав. Выделение всевозможных метаболитов и продуктов разложения в окружающую среду влияет на кислотно-щелочные и окислительно-восстановительные условия почвы и природных вод, что в итоге изменяет верхнюю часть атмосферы. Из этой области литосферы начинают извлекаться химические элементы. Они вовлекаются в водную миграцию, что и обуславливает определённый химический состав Мирового океана и осадочных горных пород.
Таким образом, несмотря на то, что общая масса живых организмов составляет очень незначительную часть массы наружных оболочек Земли (менее 0,0001%) (Литосферы, Гидросферы, Атмосферы), суммарный вклад их в биогеохимической деятельности с учётом времени их существования имеет планетарный характер.
В.И. Вернадский писал, что вопросы биогеохимии его начали интересовать в 1891 г., когда он приступил к созданию курса по генетической минералогии в Московском университете. Вероятно, в это же время стала создаваться концепция о живом веществе. В.И. Вернадский считал, что для правильной оценки геохимического значения живого вещества, необходимо, во-первых, знать средний элементарный химический состав всех организмов живого вещества, во-вторых, выразить его количественно. Кроме того, необходимо знать вес живого вещества. Этот состав и этот вес необходимо связать с весом и составом среды, в которой это (живое) вещество находится (Вернадский, 1940). В.И. Вернадский был убеждён, что биогеохимические исследования будут важны не только для решения фундаментальных научных интересов, но и необходимы для решения большинства производственных и сельскохозяйственных задач.
К сожалению, предложения В.И. Вернадского не вызвали интереса у руководителей сельского хозяйства страны, но получили поддержку геологической службы, которая стала использовать идеи Владимира Ивановича уже в середине 30-х годов прошлого столетия.
В это же время В.И. Вернадский вводит в науку термин педосфера, т.е. кора выветривания, которая является частью биосферы. В современном понимании это – почвенный слой земли. И именно в этот период Вернадский отмечает, что химический состав Мирового океана тесно связан с мобилизацией химических элементов в педосфере и с планетарным миграционным циклом почвы – воды рек – воды океана. Более того, он показал, что педосфера играет очень важную роль в газовом обмене. Важно отметить, что предположения В.И. Вернадского о циклической миграции химических элементов в системе почва – атмосфера подтвердились спустя несколько десятилетий (Вернадский, 1923, 1934, 1960).
По сути дела, в середине 30-х годов прошлого столетия В.И. Вернадский заложил все основные направления современной биогеохимии, включая продукционный и природоохранный блоки, а также, что актуально сегодня, связь биогеохимической обстановки и здоровья человека. Но, к сожалению, основные работы в области биогеохимии были развёрнуты в нашей стране лишь во второй половине XX века.
Разделы биогеохимии
Биогеохимия, как наука, за более чем полувековой период своего существования прошла значительный путь развития. Сформировались новые направления, окрепла методологическая база, накоплен громадный материал, ещё ожидающий своего обобщения. Многие теоретические положения приобрели статус принципов, законов и аксиом. Остановимся на кратком анализе основных направлений современной биогеохимии.
Глобальная биогеохимия. Это направление касается характера взаимодействия земных оболочек на уровне Земли и вопросов их эволюционного взаимодействия. В рамках этого направления глобальными задачами являются, например, проблема происхождения Земли, атмосферы и гидросферы, литосферы и криосферы, происхождения жизни, проблемы эволюции живых организмов, а также важнейшие события в жизни Земли, последствием которых явилась целая череда изменений скоростей биогеохимических циклов и их направленности.
В рамки интересов этого направления входит также последовательное изучение структурных компонентов оболочек, а также характера их взаимодействия. На современном уровне проблема взаимодействия оболочек и характер последствий сформулированы В.О. Таргульяном, (Таргульян, 1971), который в своей теории экзогенеза раскрыл важнейшие представления о трансформационном и реакционном потенциале, а также о характерном времени – периоде, за который взаимодействующие системы приходят в равновесие. Весьма ценными являются его представления о системе продуктов, чьё происхождение самым тесным образом взаимосвязано с характером и свойствами взаимодействующих оболочек.
Биогеохимия континентов. Современная биогеохимия континентов самым тесным образом связана с их происхождением. Расчёты Кларка, а затем Гольдшмидта, показавшие близость средневзвешенного состава литосферы различных континентов, косвенно подтверждают единство происхождения континентов, в частности теорию плит. В современный же период биогеохимия континентов определяется не только возрастом и степенью выветриваемости пород, но и, зачастую в первую очередь, характером и составом живого вещества. Последние сведения говорят в пользу определённых корреляций между химическим составом природных вод, дренирующих континенты, и особенностями круговорота, а точнее типологии круговорота. Особенно ярко эта взаимосвязь показана для тех континентов, где фазы выветривания пород достигли значительных границ. В этих случаях химия природных вод обусловлена теми элементами, которые не удерживаются в биологическом круговороте. Теоретически об этом писали В.И. Вернадский (1960, 1980), затем В.А. Ковда (1973,1985). Более определённо эту идею обосновал теоретически Г.В. Добровольский, который связывал систему круговорота и природных вод довольно жёстко, когда подчёркивал, что «выносятся, прежде всего, те элементы, которые не удерживаются в биологическом круговороте». Последнее положение предложено называть правилом Добровольского (Богатырев, 2015).
Биогеохимия регионов. В геологии под регионами понимают крупные территории, имеющие общие черты геологического строения. Для общей биогеохимической ситуации это имеет огромное значение. Так, А.Е. Ферсман подчёркивал, что два основных фактора определяют геохимию данной местности – её геологическое прошлое и климатическое настоящее. Климат, в свою очередь, определяет характер растительности, а соответственно и тип круговорота (Ферсман, 1955, 1958, 1959). Такими регионами, например, являются Русская платформа, Казахстанская складчатая система и др. В наиболее полной мере эта проблема решена для Русской равнины, для которой В.А. Ковда предложил гипотетическую схему её развития в послеледниковый период. Эта схема предполагает единство происхождения и генезиса осадков в пределах этой обширной территории, что определяет специфику современных процессов, в том числе и почвообразования (Ковда, 1973, 1985).
Биогеохимия биомов. Это одна из наиболее детально разработанных в биогеохимическом отношении систем. Прежде всего, этим мы обязаны блестяще разработанной классификации биологического круговорота, предложенной Н.И. Базилевич. В рамках биогеохимии это даёт принципиально важную основу для анализа особенностей биогеохимических процессов в пределах биомов (Базилевич, 1993; Базилевич и др., 1986).
Биогеохимия геохимических ландшафтов. Ядро этого направления составляет учение о геохимических ландшафтах (Полынов, 1956). Важнейшая биогеохимическая идея для геохимических ландшафтов была реализована А.И. Перельманом, который подчёркивал, что биологический круговорот является одним из важнейших законов геохимии ландшафтов. Обращая существенное внимание на значение биологической составляющей геохимического ландшафта, А.И. Перельман предложил группировку биогенных ландшафтов, основываясь на характеристиках продукционного процесса (Перельман, 1977; Перельман, Касимов, 1999). Несомненно, нельзя не упомянуть и классификацию геохимических ландшафтов по М.А. Глазовской, основанную на важнейших характеристиках растительного покрова (Глазовская, 1964).
Биогеохимия биогеоценозов. Реализация исследования биогеохимических процессов на этом уровне в своей основе связана и обусловлена использованием блестящей систематики биогеоценозов и самого этого понятия по В.Н. Сукачеву (Сукачев, 1964). Многочисленные модели разработаны для разных типов биогеоценозов.
Биогеохимия отдельных блоков биогеоценозов. Исследования на этом уровне осуществляются в зависимости от степени детальности разделения биогеоценозов на составляющие, при условии, что данная подсистема обладает определённым уровнем целостности и может быть выделена не только как компонент биогеоценоза, но и определена в точных, причем в дискретных, границах, которые обусловливают её минимальную эмерджентность. Именно на этом уровне речь может идти о биогеохимии почв, природных вод, живых организмов. При дальнейшей детализации речь может идти, например, о биогеохимии лесных подстилок, почвенно-грунтовых водах, об определённых видах растений и т.д. Великолепным примером последнего подхода являются исследования А.Л. Ковалевского, предложившего целый комплекс показателей, характеризующих именно область биогеохимии видов растений (Ковалевский, 1991).
Биогеохимия изотопов. В этом отношении хорошо известно в почвоведении использование С-12/С-14 в целях определения возраста почв. Академик И.П. Герасимов в своё время сформулировал положение о двух формах углерода. Первая форма – это та часть, которая активно вращается в круговороте, и другая часть, которая вышла из круговорота и относительно накапливается в почве. Огромное значение в настоящее время имеет использование стабильных изотопов, например кислорода, которые позволяют оценить возможные тренды, касающиеся температурных условий в палеогеографическом аспекте. В настоящее время большое значение приобрела и биогеохимия таких загрязняющих элементов, как цезий-137 и ряда других (Богатырев и др., 2007).
Историческая биогеохимия. На этом уровне выделяется, по крайней мере, два уровня исследований. В первую очередь, это весь период Земли. Второй уровень, имеющий для нас значение – это уровень голоцена, интересный в том отношении, что даёт возможность оценить изменения, обусловленные сменой биоклиматических условий. На сегодняшний день предполагается, что наиболее интенсивное почвообразование, а значит и активные биогеохимические циклы, были приурочены к субатлантическому и атлантическому периоду. Обращает на себя внимание и то, что в плейстоцене разнообразие природных зон было гораздо ниже. Их всего было 4 по сравнению с 12 биомами в настоящий период. Вместе с тем некоторые исследователи предполагают, что активное формирование почвы, по крайней мере её дифференциация, предшествовали активной роли биологической составляющей.
Биогеохимия техногенных ландшафтов. Хотя термин техногенез давно бытует в геохимии, но в последние годы значение исследований в этой области резко возросло. Это требует особых методик и особых подходов. Причём довольно часто проблемы в техногенных зонах чрезвычайно специфичны и обусловлены спецификой загрязнения и её источниками. Так, широко известны техногенные зоны около заводов, свалок, шахт и т.п., загрязнение нефтью и пр. Индикационная роль живых организмов признана чрезвычайно важной (Глазовская, 1997).
Биогеохимия специфических районов обусловлена природными аномалиями – недостатком или избытком элементов. В этом отношении фундаментальные идеи были заложены ещё В.И. Вернадским и реализованы в виде так называемых биогеохимических провинций (Вернадский, 1940, 1980). .
Экспериментальная биогеохимия. Фактически, в интегральном виде эксперимент проводится на всей планете, начиная от первого акта воздействия человека на природу. Распашка территорий, внесение удобрений, смена одних ценозов на другие, масштабные осушения и орошения громадных территорий – это все примеры изменения изначальных циклов органического вещества и элементов. Невиданный по своей значимости природный эксперимент – внесение в естественные циклы элементов радиоактивных изотопов, причём на различных по своей географии природных регионах – это Чернобыльская катастрофа. Биогеохимические методы поиска рудных месторождений образуют совершенно своеобразный раздел биогеохимии и имеют практическое значение.
Вышерассмотренная систематизация исследований биогеохимии основывается на последовательной детализации различных уровней организации земных оболочек. Вместе с тем это не исключает других подходов к изучению биогеохимии процессов и явлений. В их числе – исследование биогеохимии отдельных элементов, занимающее широкое место в научной литературе. Такой подход основывается на свойствах элемента, которые определяют его поведение в пределах биосферы. Это отнюдь не означает, что характеристики элементов остаются без внимания при других подходах, но при этом подходе они являются определяющими.
Задачи современной биогеохимии
В самом общем виде задачи биогеохимии сводятся к детальному изучению роли живых организмов. Существенно расширяется задача биогеохимии в рамках определения В.В. Ковальского, одного из крупнейших геохимиков и последователей В.И. Вернадского, определившего биогеохимию как науку о системной организованности биосферы и биогенных циклах химических элементов, в основе которых лежит эволюционное единство жизни, живого вещества и среды, определяющее закономерности биогенной миграции атомов и форм их биогенных соединений (Ковальский 1974, 1978, 1985).
Системная организация биосферы подразумевает исследование её структурно-функциональной организации, а это резко расширяет сферу исследований и включает в себя помимо изучения живых организмов и другие компоненты биосферы, а также смежные с ней оболочки, находящиеся с биосферой в активном взаимодействии. Как уже было написано выше, В.И. Вернадский считал, что для правильной оценки геохимического значения живого вещества необходимо, во-первых, знать средний элементарный химический состав всех организмов живого вещества, во-вторых, выразить его количественно. Кроме того, необходимо знать вес живого вещества. Этот состав и этот вес необходимо связать с весом и составом среды, в которой живое вещество находится (Вернадский, 1940). Не ограничившись этой триадой, В.И. Вернадский выдвинул четвертую задачу, которую он сформулировал следующим образом: «Есть ещё одно основное явление, мало охваченное научной работой и научной мыслью, для которого в данный момент нет простого и удобного числового выражения», и далее – «это явления правизны – левизны». Этим характеристикам, открытым великим Л. Пастером, В.И. Вернадский придавал исключительное значение с точки зрения фундаментального отличия косной материи и живого вещества. В настоящее время в число констант вошли Кларки, как среднее содержание во всей литосфере в целом или в её отдельных составляющих. На всем протяжении становления биогеохимии исследования велись во всех важнейших оболочках: атмосфере, литосфере, гидросфере и педосфере.
Для того, чтобы все эти задачи были корректно выполнены, специалистам различных областей необходимо правильное и единое понимание термина – биосфера. Поэтому в следующем разделе мы попытаемся уделить внимание именно этому вопросу.
Что такое биосфера
Фундаментальная концепция биосферы заключается, прежде всего, в том, что априори признается её самостоятельность в системе земных оболочек, включая специфические законы её формирования, при которых ведущее значение принадлежит живым организмам.
Эдуард Зюсс (1831-1914), профессор Венского университета, в 1875 году понимал под биосферой «область, пронизанную жизнью» и «сферу обитания живых организмов». Н.М. Сибирцев (1860-1900), российский геолог и почвовед, ученик В.В. Докучаева, ещё до основных работ В.И. Вернадского определил биосферу как особую оболочку. Однако только работами В.И. Вернадского было заложено научное представление о структурно-функциональной организации биосферы, включая её компонентный состав и специфику функционирования. В центре пристального внимания В.И.Вернадского при изложении концепции биосферы всегда лежало учение о живом веществе. В представлениях и разработках В.И. Вернадского биосферой является наружная оболочка Земли, охваченная геохимической деятельностью живого вещества. Главным отличием в таком понимании термина биосфера является то, что В.И. Вернадский воспринимал биосферу не только как место обитания живых организмов, но как глобальную систему взаимодействия живого и неживого вещества. В.И. Вернадский писал, что вопросы биогеохимии его начали интересовать в 1891 г., когда он приступил к созданию курса по генетической минералогии в Московском университете. Вероятно, в это же время стала создаваться концепция о живом веществе (цит. по Добровольский, 2003).
Многие исследователи вслед за В.И. Вернадским давали определение биосферы. Одно из удачных определений принадлежит В.А. Ковде (1972): «биосфера – это сложная многокомпонентная общепланетарная термодинамически открытая саморегулирующаяся система живого вещества и неживой материи, аккумулирующая и перераспределяющая огромные ресурсы энергии, и определяющая состав и динамику земной коры, атмосферы и гидросферы». В его определении важным является несколько аспектов. Основной аспект в определении – это системный подход и саморегулируемость биосферы, что определяет ее устойчивость.
Компоненты биосферы
В рамках концепции В.И. Вернадского в биосфере представлено три группы компонентов, генетически взаимосвязанных между собой. Первая и важнейшая из групп – это живое вещество – совокупность живых организмов. Вторая группа – это биогенное вещество (продукты, созданные живым веществом, например, угли, сапропели, гумус). Третья группа включает в себя биокосные образования – продукты, образовавшиеся в результате взаимодействия живых организмов и неживой материи (почвы, осадочные породы, некоторые газы).
Свойства биосферы
К важнейшим свойствам биосферы относят, в частности, разнообразие живых организмов, имеющее различный характер. Оно включает в себя не только размерности живых организмов, но и специфику их состава, и скорости обновления живых организмов. Не случайно в изучении этих трёх основных характеристик В.И. Вернадский видел основные задачи биогеохимии.
Другое важнейшее свойство биосферы – это ассиметричность распределения живого вещества по поверхности планеты, которая наблюдается от Арктики до тропических ландшафтов. Отметим, что неравномерность запасов живого вещества наблюдается на всех уровнях организации биосферы.
Следующее свойство биосферы – это пластичность. Это свойство в большей степени характерно, прежде всего, живому веществу, благодаря различным механизмам, включая эволюцию. Пластичность связана с другим свойством – резистентностью, относительной устойчивостью. Именно это свойство в последние годы активно исследуется в почвоведении и биогеохимии. Кроме того, как и для живых организмов, устойчивость биокосных тел также обусловливается различными механизмами – от устойчивости минералов, входящих в состав почв, до содержания гумуса и условий функционирования.
Развивая идеи В.И. Вернадского, В.А. Ковда предложил ряд новых положений относительно свойств биосферы. Им предложены такие свойства как множественность, гомеостатичность, постоянство притока и аккумуляции энергии, расширенное воспроизводство биомассы, биогеохимический круговорот, гетерогенность. Так, множественность расширяет понятие о разнообразии компонентов биосферы, а гомеостатичность представляет собой современное более широкое толкование пластичности и резистентности, хотя в строгом смысле этого слова гомеостатичность характерна исключительно для живых организмов. Постоянство притока и аккумуляции энергии в биосфере, прежде всего, обеспечивают фототрофные организмы. Фотосинтез является основным процессом в накоплении энергии на поверхности Земли. Расширенное воспроизводство биомассы – это фактически идея В.И. Вернадского о постоянно растущем выявлении живого вещества, о его стремлении занять максимально возможное пространство.
Биогеохимический круговорот отнесён к важнейшему свойству биосферы. Это свойство биосферы В.И. Вернадский называл основным механизмом функционирования биосферы. Он писал, что в биосфере существует непрерывный ток атомов, который пронизывает биосферу и определяет её существование. На современном этапе закон биологического круговорота возведён в ранг одного из основных законов геохимии ландшафта (Перельман, Касимов, 1999). В течение развития Земли эволюционировали и её оболочки, в том числе и биосфера. В соответствии с этапами изменялась и направленность циклов круговорота. Гетерогенность существенно расширяет представление о многокомпонентности, подчёркивая различие в происхождении важнейших компонентов биосферы, причём также на разных уровнях её организации. Так, гетерогенность почвы играет особую роль в распределении микроорганизмов и растительности.
Границы биосферы
Этот вопрос далеко не случайный, так как его решение имеет прямое отношение к расчёту химического состава биосферы и определению её соотношения с другими сферами. Предложено несколько решений при определении границ биосферы (Богатырев, Рыжова, 1994).
Первое – это формальный подход, при котором, по В.И. Вернадскому, мощность биосферы ограничивается несколькими километрами, охватывая не только подстилающие почву породы, но и довольно значительную часть более глубоких горизонтов, включая часть магматических пород. Современные исследования показывают, что биосфера охватывает слои литосферы, гидросферы и тропосферы суммарной мощностью около 40 км (Башкин, 2004).
Второе решение может быть названо структурным, при котором к биосфере относят области, где активность живых организмов наивысшая. Но в этом случае фактически речь идёт о части биосферы, которая, согласно А.Н. Тюрюканову (2001), получила название витасферы, а за рубежом именуется парабиосферой. В рамках структурного подхода к биосфере следует относить области с наличием всех компонентов, входящих в состав биосферы: живых организмов; продуктов их жизнедеятельности (биогенного вещества); биокосного вещества – сложного продукта взаимодействия живого вещества, биогенного и косного. В результате взаимодействия этих компонентов формируются системы, обладающие уже совершенно новыми эмерджентными, или сверхаддитивными свойствами. Примером появления таких свойств, в частности, является формирование почв с особыми качествами – это плодородие.
В основе третьего решения о границах биосферы лежит функциональный подход. В его рамках сложным вопросом остаётся мера включения в биосферу части косного вещества. Ответ на этот вопрос частично был дан В.И. Вернадским. Выше отмечалось, что в биосфере между её косной, безжизненной частью и живыми организмами идёт непрерывный ток атомов, который пронизывает всю биосферу и определяет условия её существования. Такой подход интересен тем, что биосфера в таком случае не ограничивается только тремя важнейшими компонентами, о которых речь шла выше, но в неё включается и косное вещество. Если же принимать во внимание все косное вещество, то в рамках этой концепции во многом теряется специфика биосферы. Более разумным, вероятно, необходимо учитывать только то косное вещество, которое активно участвует в процессах обмена веществом и энергией с живыми организмами (Богатырев и др.,2004).
Для системы, в пределах которой может осуществляться этот процесс, достаточно двух компонентов, с одной стороны, одним из постоянных компонентов должен быть всегда живой организм или совокупность организмов, а с другой – любые компоненты биотической, биокосной или биогенной природы. Участие тех или иных компонентов может быть ограничено.
Кроме биогенного тока, В.И. Вернадский определил другой важнейший признак, отличающий биосферу от других оболочек – пластичный эволюционный процесс, который в большей степени свойственен живому веществу.
Таким образом, в рамках функционального подхода В.И. Вернадским определён основной критерий биосферы, а следовательно, и её границ – это круговорот элементов, который можно назвать универсальным ключом к идентификации биосферы как специфической оболочки земли. В пределах биосферы поток элементов носит циклический характер.
Химический состав биосферы
Взаимодействие живой и косной систем происходит в форме массообмена химическими элементами. Как считал В.И. Вернадский, именно эти процессы поддерживают «геохимическую организованность» биосферы. Так как все эти процессы осуществляются в результате жизнедеятельности организмов, то они были названы им биогеохимическими.
Известно, что при всем многообразии живых организмов (в силу термодинамической открытости этих живых систем), общим условием их существования является обмен веществ со средой обитания. В результате своих физиологических потребностей, живые организмы поглощают химические элементы. Так как в большинстве случаев это поглощение осуществляется селективно, то это вызывает дифференциацию элементов в окружающей среде. Очень важным аспектом жизнедеятельности организмов является также их газовый обмен. Газообразные метаболиты, поступая в атмосферу, постепенно меняют ее состав. Выделение всевозможных метаболитов и продуктов разложения в окружающую среду влияет на кислотно-щелочные и окислительно-восстановительные условия почвы и природных вод. Из этой области литосферы начинают извлекаться химические элементы. Они вовлекаются в водную миграцию, что и обуславливает определенный химический состав Мирового океана и осадочных горных пород.
До сих пор определение химического состава биосферы представляется довольно сложной задачей. Существует ряд важнейших закономерностей, определяющих химический состав биосферы: как общие, присущие всем другим сферам, так и специфические. Среди них следует назвать преобладание чётных ядер над нечётными (правило Оддо-Гаркинса), а также положение о том, что логарифмическая кривая кларков является функцией заряда их атомов.
Кроме того, согласно теории Гольдшмидта, наиболее значительная роль в биосфере в силу её расположения в пределах верхних оболочек принадлежит литофильным и атмофильным элементам.
Среди специфических закономерностей следует назвать обогащённость биосферы элементами, которые присущи живым организмам. В зарубежных работах подчеркивается, что биосфера карбоксилирована и гидратирована в химическом смысле слова. Ранее также отмечалось, что в химическом составе растений, как правило, чаще всего преобладающих в биосфере (Тиссен, 1954), накапливаются те элементы, которые активно участвуют в построении первичных минералов. Согласно этим эмпирическим сопоставлениям показано, что в основном это элементы, расположенные в таблице Менделеева до 30 номера.
Несомненно, что химический состав биосферы каждый раз будет обусловлен подходом при определении её границ. На суше, при формальном выделении границ, химический состав биосферы будет определяться характером отложений при минимальном вкладе живого вещества. При отождествлении биосферы и живого вещества (в этом случае, как правило, речь идёт о растительности) состав биосферы будет целиком зависеть от набора основных эдификаторов, входящих в состав растительного покрова и слагающих данные участки биосферы. Использование функционального подхода при расчёте состава биосферы, несомненно, перспективно, при таком расчёте особенно будет велик вклад биофильных элементов.
Вопрос о химическом составе биосферы может быть решён следующим образом. Следуя положению В.И. Вернадского о том, что большая часть биосферы составляет совокупность живого вещества, биогенного и биокосного, этот комплекс и следует принять за основу при расчете химического состава биосферы. Кроме того, сюда следует отнести и все случаи биогенного минералообразования, например, некоторых осадочных пород. При таком подходе резкого преобладания в химическом составе элементов, содержащихся в изверженных и метаморфических породах, не будет, и пространственная дифференциация биосферы по химическому составу окажется более существенна.
Таким образом представляется наиболее реальным рассчитывать средневзвешенный химический состав таких ведущих компонентов биосферы, как живого, биогенного и биокосного вещества, тем более, что это может быть сделано наиболее точно по сравнению с другими подходами.
Можно сформулировать несколько положений относительно химического состава биосферы.
Первое: чем меньше вертикальная мощность рыхлых отложений, чем ближе к поверхности лежат изверженные породы, тем ближе будет состав биосферы к средневзвешенному составу живого, биогенного и биокосного вещества (пример – участки биосферы с распространением лесных экосистем на изверженных породах).
Второе: в наибольшей степени вещество биосферы будет карбоксилировано и гидратировано в химическом смысле слова, например, на территориях, с широким распространением торфов и гидроморфных ландшафтов (пример – Западная Сибирь).
Третье положение говорит о том, что в отдельных регионах распространение специфических биокосных или биогенных тел природы может оказать существенное влияние на региональный химический состав биосферы. Такая ситуация, вероятно, будет характерна для регионов с близким расположением к поверхности нефти, каменного угля, осадочных пород, или районов со специфическими видами сапропелей.
Следовательно, решение проблемы химического состава биосферы самым тесным образом связано с концептуальным подходом, прежде всего, к границам биосферы.
Обобщения, принципы, правила, законы и аксиомы современной биогеохимии
В естествознании научные исследования обобщаются различными способами, в том числе и путём установления некоторых интегративных положений, которым придаётся определенный статус. Это и интегративные обобщения, и принципы, и правила, и законы, и аксиомы. Например, в общей экологии очень удачные обобщения были сделаны Н.Ф. Реймерсом (1994). В биогеоценологии большую известность получили пять принципов, связанных с выделением биогеоценотических систем (Бяллович, 1973). В геохимии ландшафтов был выделен один из главных законов, названный законом о биологическом круговороте (Перельман, 1989; Перельман, Касимов, 1999). В почвоведение несколько базовых законов было сформулировано Соколовым (Соколов, 2002). Большую ценность имеют правила гумусообразования, предложенные Д.С. Орловым (Орлов и др., 1996). В учении о биосфере также было сформулировано значительное количество обобщений. Важно отметить, что все они не абсолютны. Согласно Рьюзу (1977), они должны быть отнесены к законам второго порядка.
Важно подчеркнуть, что обобщения происходят на различных уровнях. Первый – это уровень таких элементарных единиц биосферы, какими являются, например, почвы и живые организмы. Второй – это уровень биогеоценозов. Третий – уровень биогеоценотических систем и биогеоценотического покрова и далее биосферы.
Аксиомы. Сущность предложенных аксиом различна. Так, аксиома непрерывности подчёркивает генетическую взаимосвязь современного живого вещества с живым прошлых эпох, тогда как аксиома однородности постулирует существование живых организмов в течение всех геологических периодов. Аксиома стационарности заключается в постоянстве химического состава и общего количества живых организмов. Аксиома эргодичности свидетельствует о детерминированности процесса эволюции биосферы по отношению к начальным периодам истории её существования (Крапивин и др., 1982).
Принципы. При характеристике живых организмов одно из важнейших мест принадлежит двум принципам энергетического баланса, сформулированным Н.И. Калабуховым (1946), которые отнесены, в частности, в число фундаментальных принципов экологии. В этом контексте нельзя не упомянуть экологическое правило Одума (1986), которое систематизирует соотношение численности и биомассы живых организмов.
К числу интегративных, несомненно, следует отнести принцип максимальной замкнутости и устойчивости круговорота (Шилов, 1986), что обеспечивается взаимодействием трех фундаментальных экологических категорий – продуцентов, консументов и редуцентов на различных уровнях организации биосферы. Следует отметить, что в наибольшей степени этот принцип выражен в условиях максимального проявления живого вещества, например, в пределах витасферы, в рамках концепции А.Н. Тюрюканова (Тюрюканов, 19… ).
Особенный характер носит принцип эмерджентности (Одум, 1986), постулирующий появление новых свойств в системе и объясняющий возможность изучение целого без тщательного рассмотрения его компонентов. Таким эмерджентным свойством является, например, продуктивность. В числе теоретических следует рассматривать принцип системного анализа, имеющий широкое методологическое значение не только при изучении биологического круговорота в биосфере (Ляпунов, Титлянова,1971; Гильманов, Базилевич,1983; Титлянова, 1984), но и других сфер.
Законы. Закон биологического круговорота, возведён А.И. Перельманом (1989) в один из ведущих законов геохимии ландшафта, свидетельствует о том, что круговорот является механизмом, обеспечивающим само существование биосферы. Этот закон самым тесным образом связан с законом Вернадского, постулирующим ведущую роль живых организмов в миграции элементов. Закон Кларка – Вернадского раскрывает особенности всеобщего рассеивания элементов, причем для редких элементов это явление является преобладающим (Богатырев,Телеснина,2010).
Возможно, к числу дискуссионных положений можно отнести закон сохранения общей биомассы (Крапивин и др., 1982), показывающий постоянство количества живого вещества в течение всего геологического времени. Несомненную актуальность в современный период имеет закон Бауэра-Вернадского, выведенный в 1985 г. В.П. Казначеевым (1985), который объединяет общебиологические принципы Бауэра и биогеохимические принципы В.И. Вернадского. Этот закон фактически является характеристикой реальности, в которой происходит взаимодействие двух материальных систем – живого вещества и косной материи в единстве и противоположности. Несомненна ценность положений В.И. Вернадского о ноосфере, возведенных в ранг закона, а также закона обратимости биосферы. Последний, именуемый в некоторых работах как закон Дансеро (цит. по Дедю, 1989), в своей основе использует идеи В.И. Вернадского относительно давления жизни и стремления живых организмов занять максимально возможное пространство.
Тем не менее, как уже было сказано выше, многие законы следует рассматривать как законы второго порядка по сравнению с фундаментальными законами физики и химии, поэтому необходимо осторожно подходить к их абсолютной трактовке.
Исторически сложилось так, что многие идеи В.И. Вернадского, которые носили в его работах предположительный характер, к настоящему времени возведены в ранг законов. В связи с этим подчеркнём важность эмпирических обобщений, которые, по В.И. Вернадскому, имеют большее значение по сравнению с теориями и гипотезами. В целом же несомненная ценность подобных обобщений в виде аксиом, принципов и законов заключается в упорядочивании теоретической мысли при исследовании биосферы.
Некоторые исторические параллели развития физиологии растений и биогеохимии
В конце ХVIII в. были открыты кислород, азот и диоксид углерода. Антуан Лавуазье решил задачу количественной оценки химических элементов, участвующих в газовом обмене растений. Он пришёл к выводу об универсальности механизма круговорота при взаимодействии живых организмов с природой. Им написан трактат «Кругооборот элементов на поверхности земного шара», где он обосновал идею циклического обмена химическими элементами между тремя царствами природы: минеральным, растительным и животным. Своими работами он заложил основу современных представлений о геохимии углерода в природе.
В 1841 году два французских учёных, один из основателей органической химии Жан Батист Дюма и Жан Батист Буссенго, основатель агрохимии, путешественник и натуралист, – сформулировали идею циклического круговорота газов в системе живые организмы – атмосфера. «Все, что даёт воздух растениям, растения уступают животным, животные же возвращают воздуху; вечный круг…»
Немецкий учёный Юстус фон Либих показал, что химические элементы поступают в растения двумя путями: одни как углерод из воздуха, другие – в виде водных растворов из почвы. Он показал, что растения избирательно поглощают из почвы химические элементы. В итоге он разработал теорию минерального питания растений и положил начало изучению циклической миграции элементов в системе почва – растение – почва, получившей позже название биологического круговорота. В 1840 году в Германии была издана его знаменитая книга «Химия в приложении к земледелию и физиологии растений». Им впервые была предпринята попытка рассмотреть судьбу народов и стран в связи с нарушением естественного массообмена отдельных химических элементов.
В 80-х годах XIX века в России возникло генетическое почвоведение. Его основателем был профессор Петербургского университета Василий Васильевич Докучаев. Его работы углубили и конкретизировали представление великих химиков о деятельности живых организмов на примере почвенного покрова суши.
Идеи Д. А. Сабинина
и развитие биогеохимических исследований в морской среде
За два десятилетия до активного развития биогеохимических исследований в России и мире большой интерес к различным аспектам этой науки проявил Д.А. Сабинин. Более того, Д.А. Сабинин одним из первых проявил интерес к морю.
В архивах Института океанологии АН СССР (дело 78) имеется письмо Д.А. Сабинина директору данного института о желании принять участие в работе по проблемам продуктивности моря с научным обоснованием этих исследований и перечнем основных задач этих работ от 21 ноября 1948 года. Эта работа предварительно была названа «Продуктивность использования лучистой энергии донными водорослями». В неменьшей степени Дмитрия Анатольевича интересовало и другое направление – «Азотно-фосфатное питание донных водорослей», а в дальнейшем и исследование «Физиологические особенности продуктивности водорослей».
Эти предложения директору Института в виде отдельных статей были опубликованы учеником Д.А. Сабинина – профессором Павлом Александровичем Генкелем в книге «Дмитрий Анатольевич Сабинин, 1889-1951» (Генкель, 1980).
Помимо высших наземных растений уже в конце 40-х годов Д.А. Сабинин обратил серьезное внимание на водоросли. Более того, морские водоросли. В своих воспоминаниях о Сабинине Илья Иванович Колосов и Ольга Михайловна Трубецкова писали, что «включившись в исследования по проблеме продуктивности морей, Дмитрий Анатольевич начал изучение скорости накопления органического вещества диатомовыми водорослями на различных глубинах и изменений количества и состава пигментов у них. Он считал, что биологическая продуктивность моря определяется не только интенсивностью и качеством света и теми веществами, которые учитываются при обычных гидрохимических анализах, а именно: 1) адсорбционно-связанными питательными веществами, находящимися на коллоидальных частицах в морской воде, и 2) наличием физиологически активных органических веществ, необходимых в ничтожных количествах для роста и деления морских организмов». (И.И.Колосов, О. М. Трубецкова в кн: Д.А. Сабинин. Физиологические основы питания растений. М.Изд. АНСССР.1955. – С. 486) Для исследований, связанных со второй задачей, Д.А. Сабинин использовал микроскопические диатомовые водоросли рода Rhabdonema. Эти исследования он проводил совместно с Г.М. Чихачевой и Т. Егоровой.
Помимо микроскопических водорослей, большое внимание он уделял изучению роста и развития морских макроводорослей, в частности бурой водоросли Cystoseira barbata. Интересно отметить, что при исследовании роста и развития донных водорослей Дмитрий Анатольевич начал применять введенное им в физиологические исследования наземных растений понятие структуры урожая. Эти работы он осуществлял с ведущим альгологом страны Т.Ф. Щаповой. Впервые были получены ранее не описанные особенности роста и развития Cystoseira barbata (Сабинин, Щапова, 1954). Более того, были установлены особенности морфологического строения мелководных и глубоководных форм Cystoseira. Учитывая изменение темпов роста в различные сезоны, авторам впервые удалось определить возраст многолетних водорослей, а следовательно установить временной диапазон ассимиляции этих важных представителей живого вещества.
Фундаментальность работ Д.А. Сабинина неоспорима. Анализируя его труды можно проследить, как высказанные им идеи со временем реализовались в целые направления. Вероятно, в наибольшей степени это отразилось на развитии одного из фундаментальных направлений биогеохимии – концепции биологического круговорота. В свою очередь, биологический круговорот является одним из важнейших механизмов, обуславливающих устойчивость биосферы, включая все соподчинённые системы – от живого организма до биогеоценоза. Не случайно В.И. Вернадский писал о том, что в биосфере существует непрерывный ток атомов между живой и косной материей, который пронизывает биосферу и определяет её устойчивость.
В настоящей работе, связанной с вопросами биогеохимии и биологического круговорота, нами особенное внимание было уделено именно этим сторонам научного наследия Д.А. Сабинина. Важно подчеркнуть, что в работах Д.А. Сабинина всегда очень логично соединяются хорошо поставленный эксперимент и теоретическое обоснование. Это касается и анализа элементного состава клеточного сока, и использования радиоактивной метки для отслеживания поведения химических элементов, и изучения концентрации элементов в пасоке растений. Большое внимание уделено особой роли отдельных микроэлементов. На наш взгляд, очень интересным является изучение Д.А. Сабининым химического состава солянок. Следует отметить также роль Д.А. Сабинина как переводчика ряда крупных монографий, одна из которых посвящена роли микроэлементов в жизни растений и животных, а вторая – роли воды.
Уделяя большое внимание фундаментальным вопросам минерального питания, Д.А. Сабинин выдвинул целый ряд мыслей, идей и готовых положений:
1) Положение о круговороте веществ в растениях. Это положение получило развитие в теории круговорота минеральных элементов. Позже В.В. Снакин (1987) предложил рассматривать круговорот элементов внутри растения как биологический круговорот, в отличие от его традиционного понимания как совокупности явлений, описывающих взаимодействие растения и почвы;
2) Другое важное положение касается значения корневых систем растений, которым, по словам Д.А. Сабинина, принадлежит ведущая роль как в минеральном питании фототрофов, так и в глобальном круговороте минеральных элементов. Корневую систему он рассматривал не только как поглощающий минеральные элементы орган, но и как структуру, которая, с одной стороны, взаимодействует с окружающей средой, а с другой – взаимодействует с надземной частью растения. В связи с этим интересны замечания Д.А. Сабинина о влиянии корневых систем на особенности кислотности среды. В настоящее время к исследованиям функционирования корневых систем стали относиться более внимательно. В частности, в настоящее время проведены серьёзные исследования изменений кислотно-основных характеристик в зоне корней (Максимова и др., 2012). В результате этих исследований описана роль эктомикоризы в процессах круговорота углерода в почве (Soudzilovskaia и др., 2015).
3) Исключительно важную роль Д.А. Сабинин придавал использованию в экспериментах меченых атомов. В свою очередь, эти идеи были в полной мере реализованы при решении задач, связанных с пониманием классического круговорота элементов. Наиболее отчётливо идеи Д.А. Сабинина были воплощены в жизнь в ряде работ, касающихся рециркуляции элементов и оценки скоростей передвижения элементов в системе растения – среда. Важно напомнить, что в своих работах Д.А. Сабинин связывал фотосинтез и минеральное питание, подчёркивая при этом важность роли почвенных растворов. Более того, он обращался к роли подпахотных горизонтов.
4) Большой интерес для биогеохимии представляют работы Д.А. Сабинина, связанные с продуктивностью морских водорослей и среды их обитания. В частности, Д.А. Сабинин писал о сравнительно лучшей обеспеченности морской воды углекислым газом по сравнению с его содержанием в воздухе. Уделял внимание анализу распределения энергии, в соответствии с изменением спектрального состава солнечного света на различных глубинах. Оценивал мощность фотической зоны. Совместно с Т.Ф. Щаповой изучал рост и развитие цистозейровых водорослей.
Его отдельные мысли и идеи, приведены в таблице №1. Кроме того, в этой таблице мы попытались показать, какие работы Дмитрия Анатольевича получили дальнейшее развитие в биогеохимии.
Таблица 1. Некоторые идеи Д.А. Сабинина и их дальнейшее развитие в биогеохимии
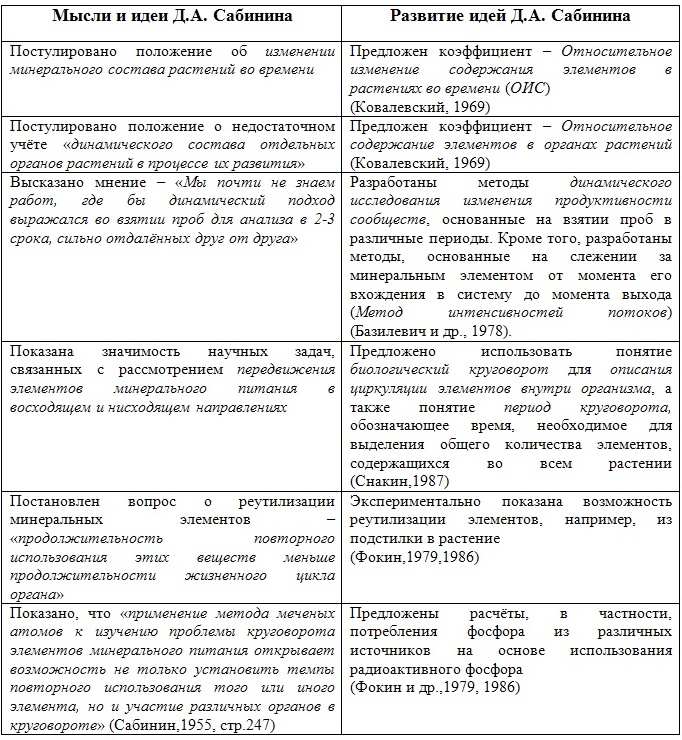
Д.А. Сабинин не только хорошо понимал роль физиологии растений для биогеохимических исследований, но и одним из первых исследователей в России начал заниматься биогеохимией Черного моря. Более того, он первым совместил эти исследования с совершенно новым направлением науки – с возрастной экологической физиологией водорослей. Ещё в середине прошлого столетия он сформировал предварительный алгоритм последовательности исследований в этой области науки. Эти исследования и подходы были высоко оценены океанологами и соратниками Дмитрия Анатольевича, но продолжение эти работы получили лишь в 70-80-е годы прошлого века.
Чёрное море как стратегически важный регион России и Европы
Как говорят мудрецы, «случайность – это почерк Бога». Вероятно, первые биогеохимические работы Д.А. Сабинина именно на Чёрном море были именно такой случайностью. Чёрное море всегда было стратегически важной областью не только для России, но и для всей Европы. Черноморский климат издавна притягивал к себе людей. Само море изобиловало рыбой, моллюсками и крупными морскими млекопитающими, что делало его мощным промысловым водоёмом. Прибрежная и водосборная зоны Чёрного моря были богаты прекрасными сельскохозяйственными землями. Выигрышное расположение между Европой и Азией делало его удобной транспортной и торговой зоной. Не исключено, что весь этот природно-климатический комплекс именно на Чёрном море способствовал прорастанию лучших традиций мировой культуры и науки.
Возраст первых стоянок морских рыболовов и охотников на побережье Чёрного моря оценивается примерно в 9 тыс. лет. С берегами Крыма связаны мифы о рождении героя Троянской войны – Ахилла. В восьмом веке до н.э. на юго-восточном побережье Чёрного моря эллины основали крупнейший центр торговли – г. Синоп. В XII-VI вв. до н.э. греческие колонии располагались не только на берегах Чёрного моря, но и в некоторых притоках Кубани, Дона, Днепра и Дуная. В те же времена здесь жили меоты, зихи, киммерийцы, скифы. Именно с Черноморским побережьем Кавказа связаны мифы о Прометее, о плавании аргонавтов за золотым руном в Колхиду. Здесь же жили амазонки.
С 30 гг. до н.э. до 64 г. до н.э. на южных берегах Чёрного моря существовало мощное процветающее Понтийское государство, возникшее в результате распада империи Александра Македонского.
Начиная с III века нашей эры в связи с опустошительными набегами и войнами, которые велись готами и гуннами, культурная и торговая значимость Чёрного моря снизилась, и лишь в VII веке, после завоевания степных черноморских просторов хазарами, значение Чёрного моря вновь возродилось. Все последующее время на черноморском побережье также велось перераспределение земель.
В качестве справки, уместно напомнить, что в разные эпохи Чёрное море имело различные названия. Наиболее древнее название – Темарун. Древние скифы называли его Ахшена, Ахщайна (др.-иранск. – темный, черный). Греки VI-V вв. до н.э. называли его Понт Аксейнос (Pontos axeinos – море негостеприимное). В начале нашей эры его, напротив, стали называть Понт Эвксинский (Pontos euxinos – море гостеприимное). Позже скифы вновь стали называть его Темным Тана. Называлось оно и Киммерийским, и Скифским, и Таврическим и, наконец, даже Русским. Сегодня оно вновь – Чёрное море (Плотников, 2007).
Сегодня по берегам Чёрного моря располагается шесть государств: с запада и севера – Болгария, Румыния, Украина и Россия, с юга и востока – Турция и Грузия. Все они активно используют ресурсы Чёрного моря.
Очень оригинальное и сложное геологическое прошлое сделало это море самым необычным морем в мире. На протяжении своей геологической истории Черное море было то пресноводным озером, то морем, т.е. имело связь со Средиземноморским бассейном. Затем вновь пресноводным озером и вновь морем и так, по крайней мере, восемь раз. 6-8 тыс. лет назад, в результате мощного землетрясения Черное море вновь соединилось со Средиземным морем. Воды моря стали солеными и произошла практически полная замена водной биоты, которая обитает в очень тонком поверхностном слое от 100 до 150 м толщиной. Остальная часть Чёрного моря «безжизненна» (Зайцев, 2006).
Площадь Чёрного моря составляет около 422 тыс. км2, объем – около 537 тыс. км3. Важно отметить, что 75% воды находится на глубине ниже 200 м, где практически отсутствует кислород. Наибольшая глубина составляет 2212 м. Площадь водосбора, т.е. суши, с которой вода стекает в Чёрное море, – 2 млн 300 тыс. км2. Это в 4,5 раза больше самого моря. На этой территории живут 160 млн человек.
Очертания Чёрного моря продолжают постоянно меняться. Оно медленно, но очень уверенно повышает свой уровень, поглощая свои берега. Осторожная оценка специалистов показывает, что за десятилетие уровень моря повышается на 3-5 см (Вершинин, 2007; Плотников, 2007).
Важно подчеркнуть, что за последние 20-30 лет Чёрное море превратилась в сильно загрязненную эвтрофированную экосистему, не способную даже поддерживать численность хищных животных высшего трофического ряда на необходимом для любой сбалансированной экосистемы уровне.
Как уже говорилось выше, Чёрное море стало, в первую очередь, очень эвтрофированной экосистемой. Прежде всего, это было связано с интенсификацией сельского хозяйства («зеленая революция»). Так, например, в северо-западной части Чёрного моря в пятидесятые годы только три большие реки в течение года вносили 13 940 т фосфатов, 154 000 т нитратов и 2 350 000 т различных органических веществ. В восьмидесятые годы вклад биогенов существенно вырос: 55 000 т фосфатов, 340 000 т нитратов, 10 488 000 т прочих органических веществ (Зайцев, 2006). Это привело к мощному увеличению количества фитопланктона (с 670 миллиграмм в одном м3 в пятидесятые годы до 30 000 в конце восьмидесятых) и, вследствие этого, количества и биомассы растительноядных видов некормового зоопланктона, в частности медузы Aurelia aurita, более чем в 300 раз. Количество кормовых видов, напротив, сократилось.
Увеличение количества планктона, включая пришельцев, привело к снижению прозрачности воды, что, в свою очередь, отрицательно сказалось на росте и развитии донных водорослей. Жесткой негативной реакцией на эвтрофирование ответила бурая водоросль Cystoseira barbata. Этот, образующий ядро биоценоза, вид цистозейры в 1970-1980 гг. просто исчез вдоль берегов северо-западного побережья. Вместе с ним исчезли 10 видов организмов, обитающих и нерестящихся в пределах таллома этого вида.
Сильное эвтрофирование привело к возникновению дефицита кислорода, или гипоксии, в придонной зоне Чёрного моря и к заморам биоты. По данным Ю. П. Зайцева, только на северо-западном участке Чёрного моря потери биомассы донных животных с 1973 по 1990 гг. составили 5 млн тонн. Количество и биомасса основных фильтраторов моря – мидий – уменьшилось на порядок (Зайцев, 2006).
Нельзя также не упомянуть о микробном, вирусном, химическом и шумовом загрязнениях, неуправляемом промысле живых ресурсов моря, негативных экологических последствиях массового туризма и рекреации.
Все это привело к снижению биологического разнообразия и полной потере некоторых черноморских видов живых организмов.
Все эти изменения являются основанием для проведения комплексных биогеохимических работ не только в водах Чёрного моря, но и в прибрежной зоне. Одним из важных направлений этих исследований должен быть мониторинг загрязнения минеральными элементами, включая как биогенные N, P, S, так и прочие элементы, которые в высоких концентрациях являются токсичными.
Характеристика состояния прибрежных вод Чёрного моря
Климат Чёрного моря смягчённый континентальный. Для территории от Таманского полуострова до г. Новороссийск характерен средиземноморский континентальный климат, от Новороссийска до Туапсе – средиземноморский континентальный климат, переходный к субтропическому, южнее же г. Туапсе климат приобретает черты субтропического. Причиной формирования этих двух различных типов климата является высота гор. До Туапсе их высота не поднимается выше одной тысячи метров, и они не являются серьёзным орографическим барьером для влагонесущих потоков воздушных масс с северо-запада. После Туапсе высота гор достигает 3 000 и более метров, поэтому на западных наветренных их склонах весь год выпадает большое количество осадков.
Температурный режим
Сезонные колебания температуры воды определяется гелиофизическими факторами и локальными характеристиками акватории (морфология дна и берегов, объем, циркуляция вод и структура гидрологических полей). Хороший летний прогрев поверхности моря обуславливает высокую (8,9°С) среднюю температуру воды. Зимой средняя температура воды на поверхности в открытом море составляет 6-8°С, однако на северо-западе и к югу от Керченского пролива опускается до 0,5°С. Летом на всей акватории моря поверхностные воды прогревается до 25°С и более до глубины 15-30 м. Глубже сезонного термоклина температура понижается примерно до слоя 75-100 м, где располагаются холодные промежуточные воды с постоянной в течение всего года температурой 7-8°С. Ниже температура с глубиной очень медленно повышается из-за геотермического притока тепла от дна и на глубине 2 км достигает 9,2°С.
Солёность
По особенностям формирования и характеристикам воды моря подразделяют на поверхностные с солёностью до 18‰, промежуточные и глубинные. Средняя солёность составляет около 18‰, близ устьев рек – менее 9‰. В открытой части моря солёность увеличивается с глубиной от 17-18‰ на поверхности до 22,3‰ у дна. Важной особенностью гидрологической структуры вод моря является существование постоянного галоклина между горизонтами 90-120 м. Солёность в этом интервале глубин увеличивается с 18,5 до 21,5‰. Сезонные изменения солёности поверхностного слоя прибрежных вод обусловливаются колебаниями соотношения речного стока и общей циркуляции. Годовой речной сток малых рек Кавказа составляет примерно 7,17 км3. Прибрежные воды от Анапы до Сочи относятся к району с относительно пониженной солёностью во все сезоны года. Стоит отметить локальное понижение солёности на юге района, в месте впадения в море рек Мзымта и Сочи. От этого участка по направлению к северу солёность повышается. Минимум в сезонном ходе 2012 года приходится на март-апрель на всех участках района и меняется от 16,39‰ (Сочи) до 17,99‰ (Анапа). Летом наблюдается незначительное повышение солёности вод прибрежья, максимум обычно отмечается в октябре-ноябре в диапазоне от 16,92‰ (Сочи) до 18,26‰ (Анапа).
Водородный показатель воды – рН
Водородный показатель воды – один из важнейших рабочих показателей качества воды, во многом определяющих характер химических и биологических процессов, происходящих в воде. В зависимости от величины pH может изменяться скорость протекания химических реакций, степень коррозионной агрессивности воды, токсичность загрязняющих веществ и т.д. Для питьевой и хозяйственно-бытовой воды оптимальным считается уровень рН в диапазоне от 6 до 9. В Чёрном море вертикальное распределение рН характеризуется наличием максимумов в поверхностном слое (0-0,5м) и на глубинах 150-200 м и локальных минимумов в слое вод 75-150 м (Коновалов, 1987).
Макрофиты как удобные организмы биогеохимического мониторинга водной среды
Одними из наиболее удобных организмов-мониторов в водной среде являются морские и пресноводные макрофиты, т.е. макроскопические водоросли, а также водные и прибрежно-водные травы. Именно те самые донные водоросли, изучением роли которых в биогеохимическом обмене азота и фосфора Чёрного моря ещё в сороковые годы прошлого века начал заниматься Д.А. Сабинин.
Водоросли – это большая группа систематически не связанных, эволюционно параллельно развивающихся талломных, бессосудистых, криптогамных, фототрофных организмов с оксигенным фотосинтезом, занимающих различные экологические ниши. Водоросли входят в различные экологические группировки: планктон, нейстон, плейстон и, наконец, бентос, а соответственно выполняют целый комплекс биосферных функций (Камнев и др., 2013)
Прежде всего, все группы водорослей являются важнейшими продуцентами морей и пресноводных водоёмов, активно аккумулирующими и переносящими химические элементы в водной среде. Кроме того, макроводоросли являются местом нереста рыб и беспозвоночных. Они укрепляют прибрежные грунты и, аккумулируя минеральные соединения, препятствуют проникновению в водную среду веществ, в частности, поступающих с береговыми стоками. Они синтезируют ценные биологически активные вещества, широко используемые в различных областях народного хозяйства.
В рамках современной систематики насчитывается 12 отделов водорослей, входящих в 9 царств, но наиболее удобными для биогеохимического мониторинга Чёрного моря являются многолетние бурые фукусовые водоросли, к которым, например, относится Cystoseira, выбранная Д.А. Сабининым (Бурдин, 1985; Камнев, 1989; Возжинская, Камнев, 1994).
Cystoseira barbata является доминирующим видом Чёрного моря. Она обитает на каменистых и каменисто-ракушечных грунтах до глубины 20 м (Зинова, 1967; Калугина-Гутник, 1975; Milchakova, 2011). Таллом C. barbata имеет моноподиальное строение. В зависимости от местообитания возраст осевой части C. barbata колеблется от 5 до 20 лет, возраст боковых ветвей – от 5 до 11 месяцев (Камнев и др., 2012).
Не менее важным и удобным объектом биогеохимического мониторинга Чёрного моря могут служить зелёные водоросли. Например, представители рода Cladophora, предпочитающие, в отличие от C. barbata, жить в самых загрязнённых эвтрофированных местах, обуславливая вторичное загрязнение этих мест (Vershinin, Kamnev, 2000). Водоросли этого рода имеют тонкое нитчатое строение. Они могут быть как прикреплёнными, так и не прикреплёнными к субстрату (Milchakova, 2011). Глубина распространения этих водорослей не превышает 20 м, а их максимальный возраст составляет всего несколько месяцев.
Среди большого количества высших водных растений можно выделить группу погруженных морских и пресноводных макрофитов, а также большую группу прибрежно-водных морских и пресноводных макрофитов.
В 1985 году И.М. Распопов (1985) ввёл термин гигрофиты, что позволило разделить все высшие водные растения на две большие эколого-физиологические группы: гидрофиты и гигрофиты. Гидрофиты – настоящие водные растения, постоянно обитающие в воде и весь цикл развития которых также происходит в водной среде. Гигрофиты – наземные растения влажных, переувлажнённых и периодически затапливаемых местообитаний с высокой влажностью воздуха. В большинстве случаев таким местом является береговая зона водных объектов или зона штормового заплеска – супралитораль.
Такое разделение способствовало более активному развитию экологии и физиологии прибрежно-водной высшей растительности. Это, с одной стороны, значительно расширило диапазон эколого-физиологических исследований прибрежно-водных растений, а с другой – сделало работу более корректной.
В пределах группы гидрофитов, в зависимости от местообитания растений, можно чётко выделить три подгруппы:
1. Подгруппу истинных гидрофитов – эугидрофитов, или гидатофитов, у которых основная масса растения находится под водой, жизненный цикл которых также проходит под водой. У некоторых их этих растений только генеративные побеги возвышаются над водой. При этом одни из них, имеющие корни, хорошо укоренены. Другие, как мхи, не имеющие корневой системы, прочно прикреплены к подводному субстрату. Наконец, третьи свободно плавают в водной толще, при этом не имеют корневой системы, например роголистник погруженный (Ceratophyllum demestrum L.);
2. Вторая подгруппа – плейстогидрофиты (или по-другому – плейстофиты) – нимфеиды, плавающие растения. Это группа водных растений с плавающими на воде генеративными и ассимиляционными органами. Среди этих растений также встречаются как укоренённые, так и неукоренённые формы;
3. Третья подгруппа – аэрогидрофиты, гелофиты, воздушно-водные растения, полупогруженные растения, болотно-водные растения. Это водные растения с побегами, часть которых находится в водной среде, а часть возвышается над поверхностью воды. Большинство растений этой группы занимает в водоёмах доминирующее положение. Так как строение стеблей и листьев этих растений приспособлено к сильному ветровому и волновому воздействию, иногда эту группу называют жёсткой растительностью.
Необходимо отметить, что в случае пересыхания или обмеления водоёма некоторые водные растения-гидрофиты (например, горец земноводный Polygonum amphibium L.) образуют наземную прямостоящую форму, не отличающуюся от обычных наземных растений.
В группе гигрофитов также можно выделить 3 подгруппы:
1. Эугигрофиты – наземные околоводные растения, приспособленные к обитанию в береговой полосе водоёмов, характерных для низких и средних уровней береговой зоны затопления. Эти растения часто встречаются в руслах неглубоких рек и ручьёв, на сплавнинах, сырых прибрежных отмелях (при глубине до 20-40 см);
2. Гигрогелиофиты – наземные болотные растения, приспособленные к обитанию в сильно переувлажнённых и даже обводнённых местообитаниях, однако нередко имеющие ксероморфное строение;
3. Гигромезофиты – наземные растения достаточно широкой экологической амплитуды по отношению к воздушному увлажнению, занимающие высокие уровни береговой зоны затопления, сыроватые или влажные отмели и зону заплеска водоёмов.
В 1988 г. Л.Ф. Лукина и Н.Н. Смирнова (1988), используя терминологию Распопова И.М., сохранив исходный смысл классификации Б.А. Федченко (1949) и А.П. Шенникова (1950), предложили для экологов и физиологов растений следующую схему подразделения высших водных растений на 5 экологических группировок:
1. Гелофиты.
2. Укорененные плейстофиты.
3. Укорененные гидатофиты.
4. Неукорененные или свободноплавающие плейстофиты.
5. Неукорененные гидатофиты.
Важно подчеркнуть, что данный подход может быть очень удобным при решении многих биогеохимических задач.
С точки зрения гидробиологической терминологии, высшие водные растения могут входить в группировку бентосных, планктонных и плейстонных сообществ.
Среди большого количества морских высших растений наиболее важными считаются морские травы (seagrasses), которые являются реальными гидрофитами и обитают в различных морях. Все морские травы принадлежат к классу однодольных (Lipiopsida), порядкам наядовых (Najadales) и частуховых (Alismatales).
Наядовые, обитающие в морской среде, включают 10 родов. Роды – Zostera, Phyllospadix, Heterozostera, Posidonia, Amphibolis, Ruppia обитают в морях умеренных и северных широт. Роды Cymodoсea, Syringodium, Thalassodendron – в тропических водах Важно отметить, что отдельные виды Ruppia способны жить не только в морях и солёных озёрах, но и в пресных водах.
Важными достаточно широко распространёнными водными растениями являются рдесты (род Potamogeton). Их насчитывается более 100 видов. Они встречаются в солёных и пресных водах умеренных широт обоих полушарий.
Морские частуховые растения обитают преимущественно в тропических регионах различных континентов, а также умеренных водах Африки и Австралии. В начале прошлого столетия они стали интродуцентами Средиземного моря.
Наиболее значимыми морскими представителями этого порядка являются: талассия, по-другому черепашья трава (Thalassia), солелюбка (Holophila) и энгалус (Enhalus). Все они относятся к семейству водокрасовых (Hydrocharitaceae).
Помимо истинных типичных морских трав в литоральной зоне и на солёных маршах обитают и другие представители однодольных и двудольных растений. Правда, эти растения могут быть затопленными только во время приливов. Наиболее известными из них являются злаки семейства Poaceae, образующие заросли на солёных маршах. Среди двудольных галофильных растений разнообразие достаточно широкое. В затапливаемой литоральной зоне живут представители семейств первоцветных (Primulaceae), маревых (Chenopodiaceae), сложноцветных (Compositae), ситниковых (Juncaginaceae), подорожниковых (Plantaginaceae) и многие другие.
К морским черноморским травам-макрофитам относится несколько видов из 5 родов высших водных растений, обитающих преимущественно на мягких грунтах в мелководной зоне, до глубины около 7-10 м: Zostera, Stuckenia, Zannichellia, Ruppia, Potamogeton. Все они могут быть удобными объектами мониторинга.
Среди пресноводных высших растений это мохообразные (Bryophyta), включающие печёночные (Hepaticae) и листостебельные (Musci) мхи; плауновидные (Lycopodiophyta), представленные группой полушниковых (Isoetales); хвощевидные (Equisetophyta), наконец, это цветковые растения (Anthophyta), которые насчитывают большое количество видов, принадлежащих более чем к тридцати семействам. Наиболее известными из них являются ряска, кувшинка, стрелолист, водокрас, рогоз, камыш, тростник, любимые аквариумистами валлиснерия и «водяная чума» элодея.
Достаточно интересными объектами биогеохимического мониторинга являются высшие водные растения, растущие в устьях рек, впадающих в море. Эти растения, испытывая нагрузку с постоянным изменением солёности воды, также аккумулируют минеральные элементы, включая их в биохимические циклы.
На наш взгляд, из высших наземных водных макрофитов одним из наиболее удобных объектов для биогеохимического мониторинга черноморского побережья может быть использован тростник Phragmites australis (Cav.) Trin. ex Steud, который растёт вдоль берегов малых рек и заиленных заливов самого моря.
Особенности концентрирования минеральных элементов водными макрофитами
Очень важной задачей биогеохимического мониторинга является оценка диапазона аккумулятивных возможностей объектов мониторов. Выбор макрофитов в качестве модельных объектов для биогеохимических исследований, как это было написано выше, связан с их важнейшим значением для гидросферы как одних из основных первичных продуцентов шельфовой зоны Мирового океана, где их продукция сопоставима с продукцией лесов умеренного пояса. Не менее значимыми продуцентами являются и прибрежные растительные сообщества, окружающие как береговую зону морей, так и рек, несущих свои воды в эти моря. Кроме того, водные и прибрежно-водные сообщества макрофитов являются естественным барьером на пути антропогенных загрязнений, поступающих с береговыми стоками.
Для того, чтобы правильно оценить биогеохимическую обстановку региона, исследователи обычно выбирают (в зависимости от поставленной задачи) различные доминантные, но типичные представители морских макроводорослей или высших растений исследуемого района. Выбираются определённые типичные участки, в пределах которых достаточно удобно периодически (в разные климатические сезоны) отбирать пробы растительности, донных отложений, почвы и воды. Кроме того, в зависимости от возможностей лаборатории, могут быть использованы различные методы оценки минерального состава. Для этих целей чаще всего используют метод атомно-абсорбционной спектроскопии (ААС), метод рентгенофлуоресцентого анализа (РФА) или метод нейтронной активации (МНА).
В настоящей работе для демонстрации возможностей различных методов и подходов, а также для примера морфологического и экологического разнообразия фоторофов, нами были выбраны четыре представителя водных и прибрежно-водных макрофитов, принимающие на себя первый удар поступающих с береговым стоком загрязнителей и обитающие в различных экологических условиях.
1. Cystoseira barbata (Stachouse) C.Agardh – крупная (до 1м высоты) прикреплённая морская бурая водоросль, обитающая на твёрдом субстрате (каменистые террасы, валуны, камни). Поглощает всей (очень сложно дифференцированной) поверхностью таллома. Корневая система отсутствует.
2. Cladophora sericea (Hudson) Kutzing – мелкая неприкреплённая (нитчатая) морская зелёная водоросль, обитающая на песчаном субстрате, в прибрежной зоне песчаных пляжей. Поглощает всей поверхностью таллома. Корневая система отсутствует.
3. Zostera marina Linnaeus – крупное морское высшее цветковое растение, обитающее на песчаных и заиленных грунтах. (Имеет хорошо развитую корневую систему). Поглощает корневой системой и подводными листьями.
4. Phragmites australis (Cav.) Trin. Ex Steud – прибрежно-водное высшее растение, обитающее вдоль береговой зоны рек и морей. (Корневая система составляет более 80% биомассы всего растения). Поглощает придаточными корнями из почвы, донных отложений и воды.
В таблицах 2-5 приведены данные изменения содержания минеральных элементов в различных водных макрофитах, полученные нами не только разными аналитическими способами, но и с разной выборкой, полученной как в течение многолетних наблюдений (Cystoseira barbata), сезонных наблюдений, включающих нескольких станций (Phragmites australis), так и единоразового сбора (Cladophora sericea, Zostera marina). Эта небольшая иллюстрация даёт возможность наглядно продемонстрировать, что с одной стороны, изменение минерального состава различных макрофитов может быть хорошим дополнением в понимании биогеохимического состояния исследуемого региона, а с другой, – что единоразовый сбор исследуемого материала не показывает диапазона концентрационных возможностей макрофитов, что в свою очередь, может приводить к некорректным выводам. Поэтому, более правильно вести многолетний биогеохимический мониторинг, учитывая глубинные, сезонные и многолетние изменения в исследуемых объектах.
Так, в табл. 2 приведены многолетние (более 20 лет) данные изменения минерального состава Cystoseira barbata (Bunkova et al, 2014; Kamnev et al, 2014). Более того, в этой таблице собраны данные, полученные в результате многолетних исследований с применением метода атомно-абсорбционной спектроскопии (Na, K, Mg, Ca, Fe, Mn, Cu, Zn, Pb, Ni), и в течение нескольких лет, методом нейтронной активации (остальные элементы). Только такие многолетние данные позволяют относительно корректно судить о реальных концентрационных возможностях Cystoseira barbata (Stachouse) C.Agardh
Таблица 2. Диапазон изменений содержания минеральных элементов в талломах Cystoseira barbata
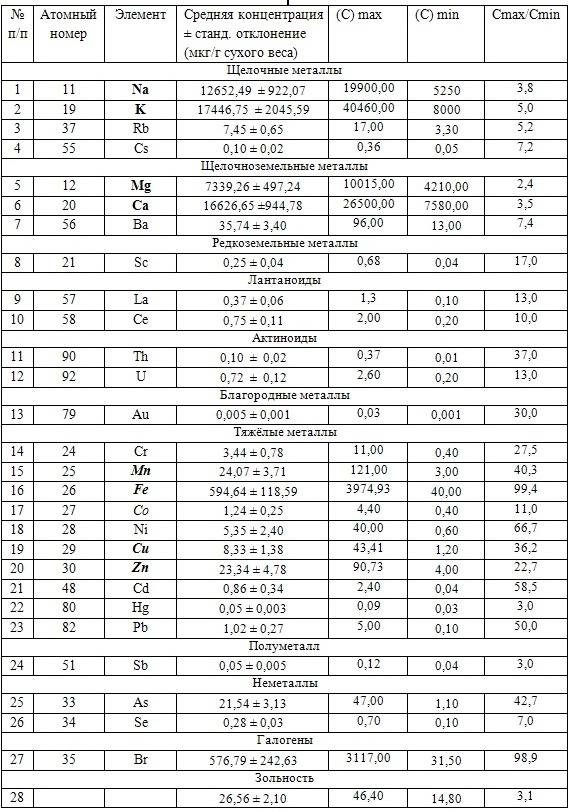
Из таблицы 2 видно, что минеральные элементы по их среднему содержанию в талломах цистозейры можно расположить в следующей убывающей последовательности: K > Ca > Br > Na > Fe > Mg > Ba > As > Mn > Zn > Ni > Rb > Cu > Cr > Co > Pb > U > Ce > La > Se > Sc > Cs > Sb > Th > Hg > Au.
Как видно, содержания одних и тех же минеральных элементов в одновозрастных водорослях в зависимости от различных факторов могут изменяться от 2 до 99 раз (в зависимости от элемента). Причиной таких колебаний может быть как изменяющаяся концентрация самого отдельного минерального элемента в воде, а также комплекса экологических факторов среды обитания, влияющего на накопление этого элемента растительным организмом, так и физиологическое состояние самого макрофита.
Соотношение максимальной и минимальной концентраций макроэлементов, например, таких как Na, K, Mg, Ca в талломе Cystoseira barbata может изменяться от 2 (Mg) до 5 (K) раз, а микроэлементов – от 22 (Zn) до 99 (Fe) раз. Аналогичные флуктуации характерны и для физиологически «незначимых» элементов. Эти результаты, с одной стороны, в какой-то степени, показывают физиологическую избирательность поглощения химических элементов Cystoseira barbata и подтверждают наличие физиологических регуляторных механизмов накопления биогенных элементов. Возможно, это обусловлено тем, что доступность этих элементов в разные геологические эпохи изменялась и, соответственно, физиологические механизмы накопления имеют разные диапазоны. С другой стороны, эти результаты показывают на наличие определённого пула, способного аккумулировать эти элементы. В случае с цистозейрой – таким накопительным пулом минеральных элементов могут служить альгиновые кислоты и фукоиданы, формирующие клеточную стенку и межклетники, и задерживающие проникновение избыточных концентраций элементов в клетку.
В таблице 3 приведены данные минерального состава зелёной водоросли Cladophora sericea (Hudson) Kutzing, собранной в августе 2013 г. в черте г. Анапа. В этом случае нами был использован метод нейтронной активации, позволяющий определить единоразово 41 химический элемент. Удобство этого метода бесспорно, но применение его не всегда возможно из-за дороговизны исследований.
Таблица 3. Диапазон изменений содержания минеральных элементов в талломах Cladophora sericea.
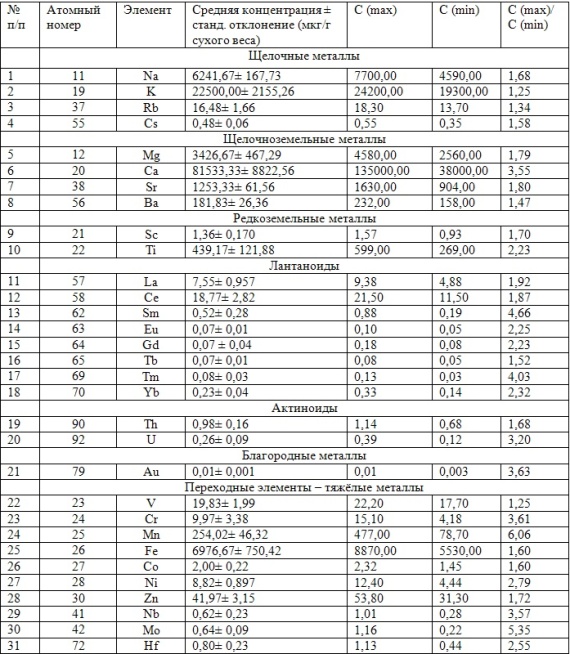
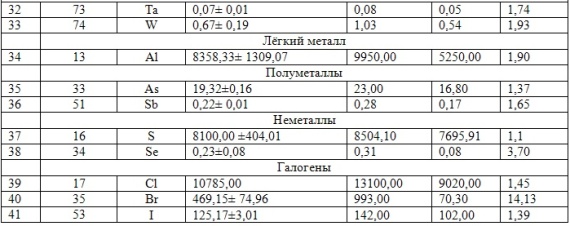
Из полученных данных видно, что минеральные элементы по их среднему содержанию в талломах Cladophora sericea можно расположить в следующей убывающей последовательности: Ca > K > Cl > Al > S > Fe > Na >Mg > Sr >Br > Ti > Mn > Ba > I > Zn > V > As > Ce >Rb >Cr > Ni >La > Co > Sc > Th > Hf >W > Mo > Nb > Sm > Cs > U > Yb >Se > Sb > Tm > Gd > Eu >Tb ≈ Ta > Au.
Интересно отметить, что в отличие от цистозейры таких серьёзных изменений в соотношении максимальных и минимальных концентраций не наблюдается. Хотя для некоторых элементов, например, таких как Mn, Mo, Br диапазон концентраций может изменяться в 5-14 раз. С одной стороны, это можно объяснить тем, что пробы брались в одном месте и в одно и то же время (т.е. не было сезонных и прочих изменений), а с другой – совершенно иной (в химическом отношении) клеточной стенкой и межклетниками у зелёных макроводорослей, а соответственно другими механизмами аккумулирования.
В таблице 4 приведены данные по изменению минерального состава в морском, полностью погруженном высшем растении – Zostera marina Linnaeus. В отличие от водорослей у высших морских гидрофитов имеется хорошо развитая корневая система, благодаря которой они укрепляются в мягких морских грунтах и поглощают минеральные элементы, как самой корневой системой из донных отложений, так и листьями. Пробы брались в августе 2012 г. в Таманском заливе, недалеко от посёлка Сенной. В этом случае нами был использован широко распространённый и достаточно удобный метод атомно-абсорбционной спектроскопии. Недостатком этого метода может считаться уменьшение количественного спектра самих минеральных элементов, хотя для многих задач этого вполне достаточно. Более того, зная регрессионные зависимости содержания одного химического элемента от другого, можно рассчитать его содержание.
Таблица 4. Диапазон изменений содержания минеральных элементов
в надземной и подземной частях Zostera marina
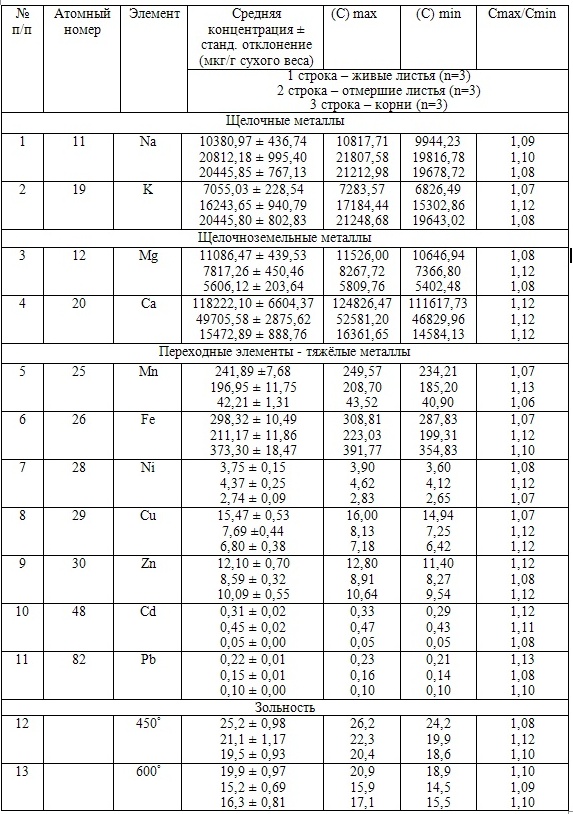
Как видно, содержание изучаемых минеральных элементов в различных морфологических частях зостеры может различаться. Концентрации минеральных элементов в различных частях Zostera marina можно расположить в следующих убывающих последовательностях:
в живых листьях – Ca > Mg > Na > K > Fe> Mn > Cu > Zn > Ni > Cd > Pb
в отмерших листьях – Ca > Na > K > Mg > Fe > Mn > Zn > Cu > Ni > Cd > Pb
в корнях – K ≈ Na > Ca > Mg > Fe > Mn > Zn > Cu > Ni > Pb > Cd
Из таблицы видно, что соотношение максимальной и минимальной концентраций проанализированных нами минеральных элементов в Zostera marina колеблется незначительно – от 1,06 до 1,13 раз, что можно, как и в случае с Cladophora sericea, объяснить незначительной выборкой растений, более того собранных в одном месте и в одно время. Приводя в пример данные, полученные разными способами, мы хотели показать, что они могут существенно различаться, но для ряда задач могут быть вполне интересны.
При сравнении содержания макроэлементов в живых, отмерших листьях и корнях, видно, что K и Na достоверно больше в корнях, а Mg и Ca, напротив, в живых листьях, что вполне объяснимо чисто физиологическими потребностями в этих элементах для различных морфологических частей растений. Кроме того, повышенное содержание Ca (его больше, практически в 8 раз) может быть связано с тем, что листья могли быть просто плохо очищены от кальцинированных обрастаний. (Этот пример приведён специально, т.к. такое часто встречается в полевых условиях). Содержание микроэлементов распределено следующим образом: Fe несколько больше в корнях, а Mn и Cu в листьях. Содержание Zn везде примерно одинаково. Концентрация токсичных тяжёлых металлов, таких как Ni, Pb и Cd существенно выше в листьях. Не исключено, что это может быть связано с повышенных содержанием этих элементов в водной среде.
Интересно отметить, что общая минерализация (об этом говорят данные о зольности) выше в листьях. Это также, в определённой степени, может свидетельствовать о том, что листья участвуют в самостоятельном процессе поглощения минеральных элементов.
В таблице 5 представлены данные о диапазоне изменений минеральных элементов в тростнике – Phragmites australis (Cav.) Trin. Ex Steud. Тростник является широко распространённым прибрежно-водным гелофитным или гигрофитным высшим растением, обитающим вдоль береговой зоны рек и морей. Его многолетняя корневая система составляет более 80% биомассы всего растения. Помимо поглощения минеральных элементов основными корнями, хорошо укоренившимися в донных отложениях или прибрежных почвах, большую роль в минеральном обмене играют придаточные корни. Они могут поглощать минеральные элементы из почвы, донных отложений и воды. Пробы были собраны в августе 2013 г. вдоль большого участка черноморского побережья от Таманского полуостров до г. Туапсе. Минеральный состав определяли методом рентгенофлуоресцентого анализа (РФА).
Таблица 5. Диапазон изменений содержания минеральных элементов
в надземной и подземной частях Phragmites australis.
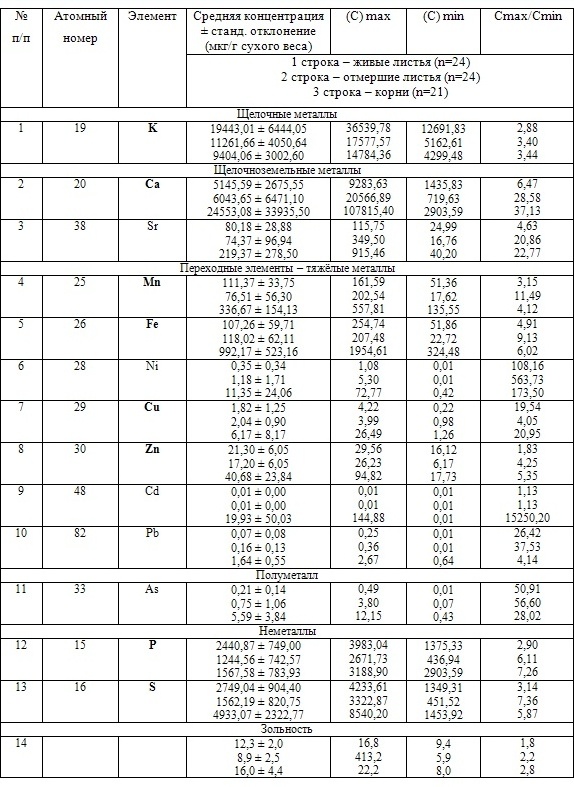
Как видно из таблицы, содержание изучаемых минеральных элементов в различных морфологических частях тростника различается. Концентрации минеральных элементов в различных частях растения можно расположить в следующих рядах убывающих последовательностей:
в живых листьях – K > Ca > S > P > Mn > Fe > Sr > Zn > Cu > Ni > As > Pb > Cd
в отмерших листьях – K > Ca > S > P > Fe > Mn > Sr > Zn > Cu > Ni > As > Pb > Cd
в корнях – Ca > K > S > P > Fe > Mn > Sr > Zn > Cd > Ni > Cu > As > Pb
При сравнении содержания макроэлементов в живых и отмерших листьях, а также корнях видно, что содержание Ca и S достоверно больше в корнях; Nа, напротив, больше в листьях (как в живых, так и в отмерших), а фосфора – в живых листьях. Содержание микроэлементов, полуметалла мышьяка и токсичных тяжёлых металлов, таких как Ni, Pb и Cd, выше всего в корнях растений. Например, содержания Fe в корнях по сравнению с листьями выше примерно в 9 раз, а Ni и Pb – более чем в 10 раз.
Интересно отметить, что самое большое различие в содержании между листьями и корнями наблюдается для Cd (15000). Это связано с тем, что часть проб была взята в очень загрязнённом (по Cd) районе, а следовательно содержание этого элемента в корнях растений, собранных из этого региона, было очень высоким и изменяло величину средних показателей. Эти результаты показывают, с одной стороны, широту аккумулятивных возможностей корневой системы гидрофитов, в частности, тростника, а с другой то, что только при увеличении выборки (хотя бы количество районов) можно оценить реальный диапазон аккумулятивных возможностей растений. Интересно отметить, что, как показывают данные о зольности, общая минерализация Phragmites australis, в отличие от Zostera marina, выше в корнях.
Полученные нами данные могут служить наглядным примером того, какие методы (в зависимости от экономических возможностей научных подразделений) могут быть использованы для оценки минерального состава водных фототрофов. Важно подчеркнуть, что при использовании различных методов, данные могут отличаться на 10-15%. Кроме того, представленные нами данные показывают, что даже минимальное изменение выборки может существенно влиять на оценку диапазона аккумулятивных возможностей растений. Для ряда задач это вполне допустимо, а для ряда – нет. Наконец, представленные нами результаты хорошо демонстрируют экологическое и видовое разнообразие физиологических механизмов накопления минеральных элементов из окружающей среды. Так, например, у полностью погруженных гидрофитов (как, например, у Zostera marina), листья также участвуют в поглощении минеральных элементов. И если их содержание в водной среде выше, чем в донных отложениях или находится в более удобной для поглощения форме, то концентрация этих элементов в листьях может быть значительно выше, чем в корневой системе.
Таким образом, все сказанное выше даёт представление том, что необходимо учитывать при постановке задач с использованием биогеохимического мониторинга.
Коэффициенты концентрирования и биологического поглощения
Одним из важнейших направлений в биогеохимии является концепция биогеохимических провинций, в основе разделения которых лежит специфика поведения элементов в пределах определённых территорий, связанная с избыточным содержанием элементов или их дефицитом в окружающей среде. Поэтому установление содержания элемента по отношению к определённому стандарту является одним из важнейших приёмов в биогеохимии (Богатырев, 2008).
Существуют подходы интегрального рассмотрения данных по содержанию важнейших макро- и микроэлементов. К таким подходам относится построение геохимических спектров элементов. При их построении руководствуются следующей методикой. Обычно на оси абсцисс последовательно помещают элементы согласно их порядковому номеру. На оси ординат откладывают отношение содержания каждого элемента в исследуемом объекте к среднему содержанию в литосфере или земной коре (кларку). Когда показатели больше 1, говорят о концентрировании элемента, меньше – о рассеивании (Богатырев, 2008).
При антропогенном загрязнении окружающей среды опасным является не только превышение ПДК элементов (предельно допустимые концентрации), но и изменение соотношений элементов друг с другом, и это следует учитывать при проведении подобного рода работ. Для расчета коэффициента концентрирования были использованы кларковые значения, установленные А.П. Виноградовым (Виноградов, 1962).
Таблица 6. Кларковые значения некоторых химических элементов в земной коре
(Виноградов, 1962)
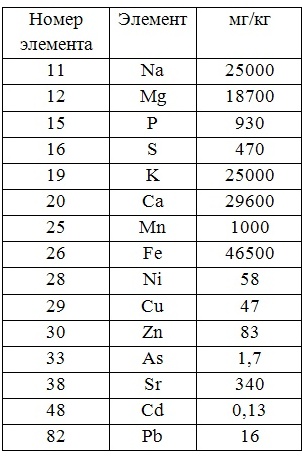
Таблица 7. Коэффициенты концентрирования и биологического поглощения элементов
в Cystoseira barbata
(м. Большой Утриш, м. Кадош, 2013 г., n=6,
метод определения элементов – рентгенофлуоресцентный анализ) *
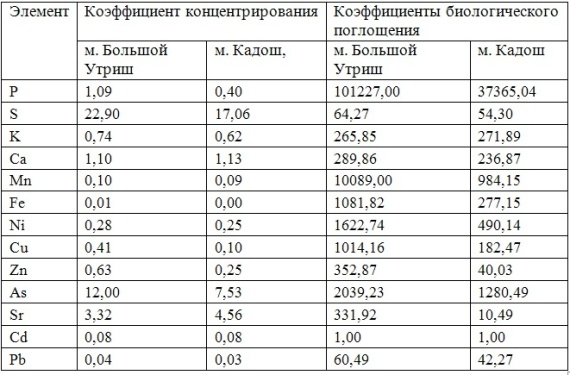
*рассчитаны на абсолютно сухую массу
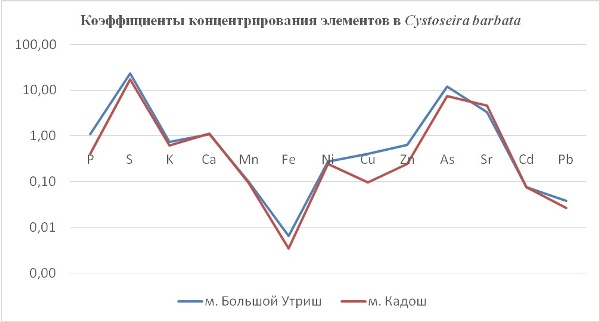
Рис. 1. Коэффициенты концентрирования элементов в Cystoseira barbata.
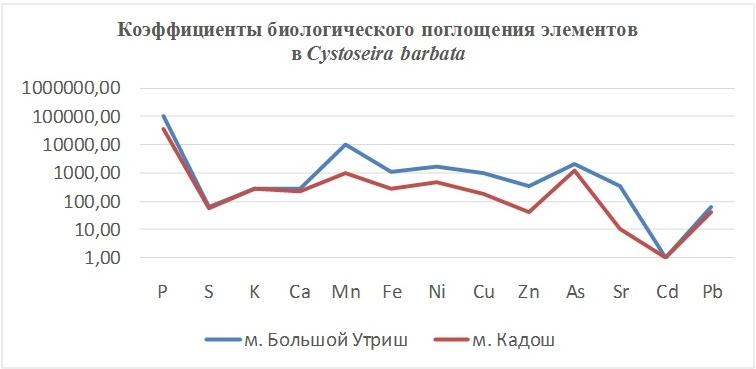
Рис. 2. Коэффициенты биологического поглощения Cystoseira barbata.
Таблица 8. Коэффициенты концентрирования и биологического поглощения элементов
в Cladophora sericea
(Анапский район, 2013 г., n=12,
метод определения элементов – рентгенофлуоресцентный анализ) *
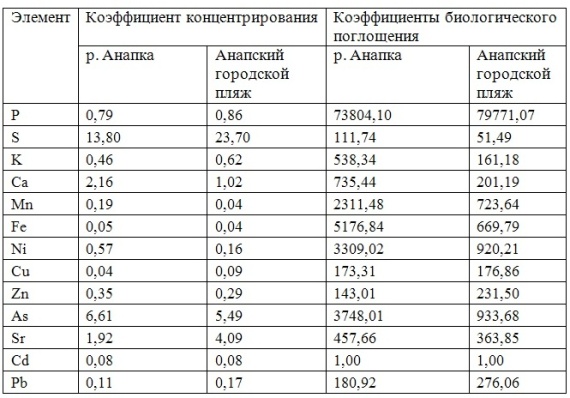
*рассчитаны на абсолютно сухую массу
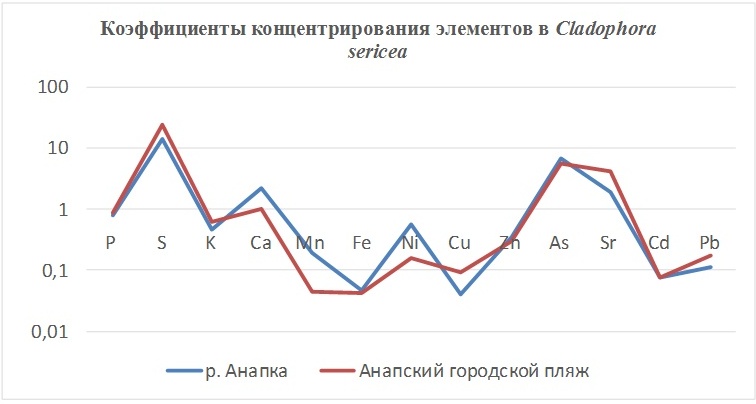
Рис. 3. Коэффициенты концентрирования элементов в Cladophora sericea.
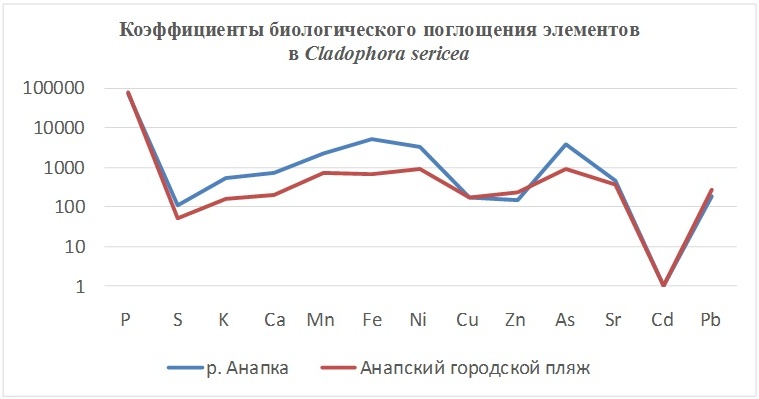
Рис. 4. Коэффициенты биологического поглощения элементов в Cladophora sericea.
На примере демонстрации геохимических спектров, представленных выше, хорошо видно, что полученные в результате исследований данные показывают графическую однотипность поведения элементов, аккумулированных в макрофитах, собранных в одном регионе. Интересно отметить, что это наглядно видно на примере графиков концентрирования и поглощения у совершенно разных групп морских водорослей (бурой макроводоросли Cystoseira barbata и зеленой макроводоросли Cladophora sericea), собранных в двенадцати километрах друг от друга. На графиках чётко выделяются одни и те же пики концентрирования серы, мышьяка и стронция. Более того, эта же тенденция поведения элементов наблюдается и у Cladophora, собранной в реке, которая впадает в море. Скорее всего, эти данные могут свидетельствовать о повышенном содержании S, As и Sr в анапском регионе в целом. Коэффициенты биологического поглощения указывают на более высокие значения этих параметров в реке по сравнению с морем. Не исключено, что это может свидетельствовать о том, что содержание этих элементов в пресной воде несколько выше и они могут находиться в другой химической форме.
Таблица 9. Коэффициенты концентрирования и биологического поглощения элементов
в Zostera marina
(пос. Сенной, 2012 г., n=3,
метод определения элементов – атомно-абсорбционная спектрометрия) *
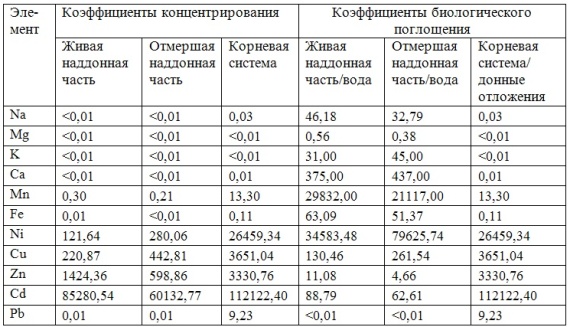
*рассчитаны на абсолютно сухую массу
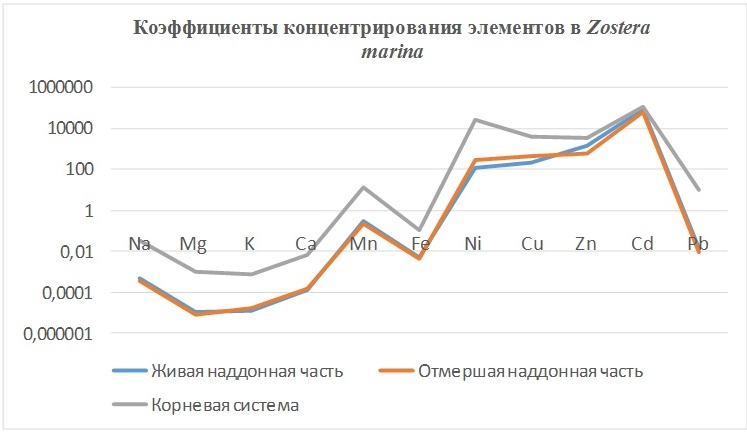
Рис.5. Коэффициенты концентрирования элементов в Zostera marina.
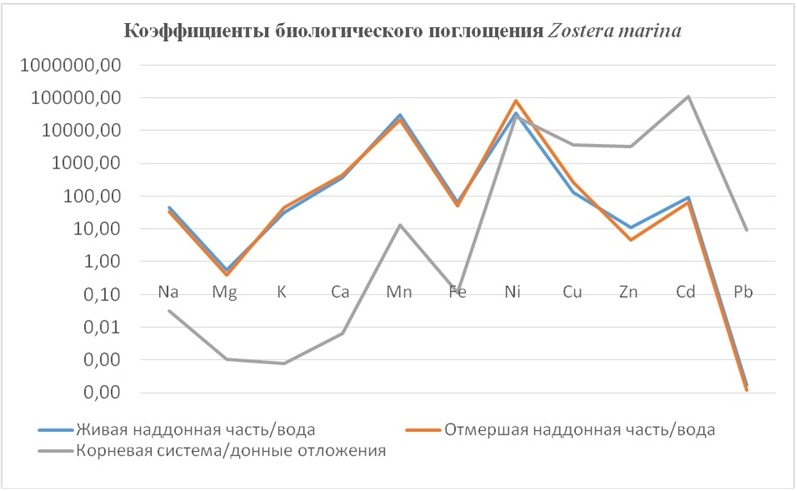
Рис. 6. Коэффициенты биологического поглощения Zostera marina.
При анализе концентрирования элементов в Zostera marina, собранной в Таманском заливе, который располагается в ста километрах севернее Анапы, нас в большей степени интересовало изменение содержания определённых тяжёлые металлов. Поэтому именно изменению этих элементов нами было уделено основное внимание. На графиках, полученных в результате исследований и представленных выше, чётко выделяются пики коэффициентов концентрирования Ni, Cu, Zn, Cd, Pb. Это, с одной стороны, свидетельствует о загрязнении данного района, а с другой – показывает, что основная роль в этом процессе принадлежит многолетней корневой системе.
Таблица 10. Коэффициенты концентрирования элементов
в отмершей наддонной части Phragmites australis
(Анапский район, 2013 г., n=18,
метод определения элементов – рентгенофлуоресцентный анализ) *
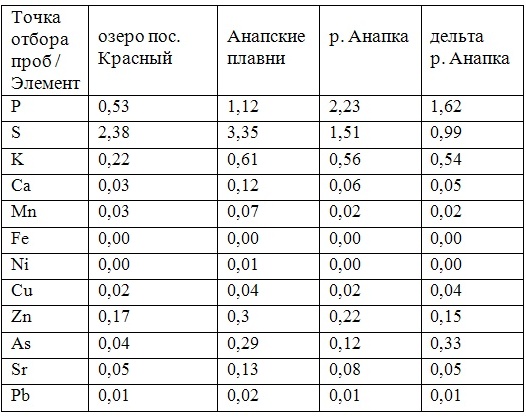
*рассчитаны на абсолютно сухую массу
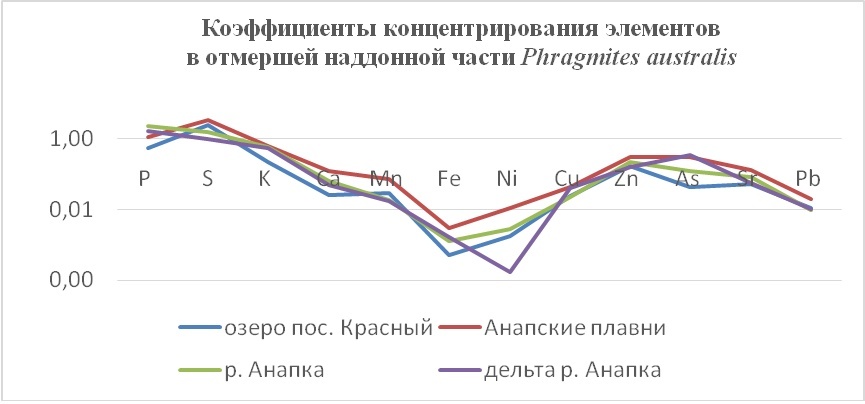
Рис. 7. Коэффициенты концентрирования элементов в отмершей наддонной части Phragmites australis.
Таблица 11. Коэффициенты концентрирования элементов
в живой наддонной части Phragmites australis
(Анапский район, 2013 г., n=18,
метод определения элементов – рентгенофлуоресцентный анализ) *
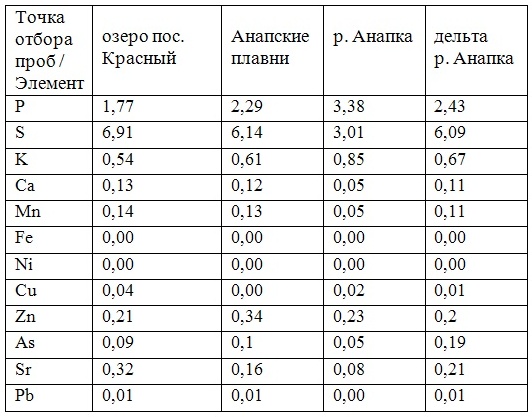 *рассчитаны на абсолютно сухую массу *рассчитаны на абсолютно сухую массу
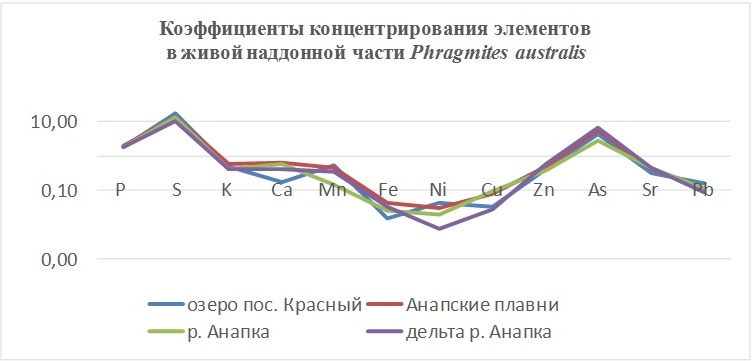
Рис. 8. Коэффициенты концентрирования элементов в живой наддонной части Phragmites australis.
Таблица 12. Коэффициенты концентрирования элементов
в корневой системе Phragmites australis
(Анапский район, 2013 г., n=18,
метод определения элементов – рентгенофлуоресцентный анализ) *
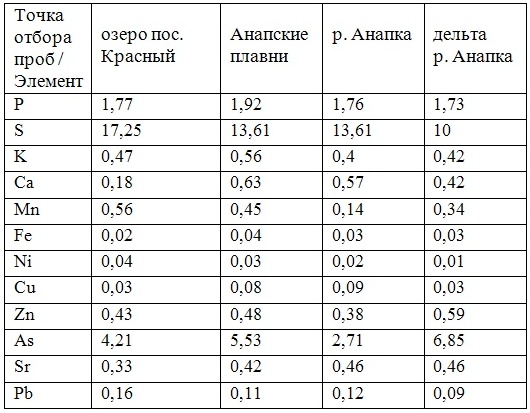
*рассчитаны на абсолютно сухую массу
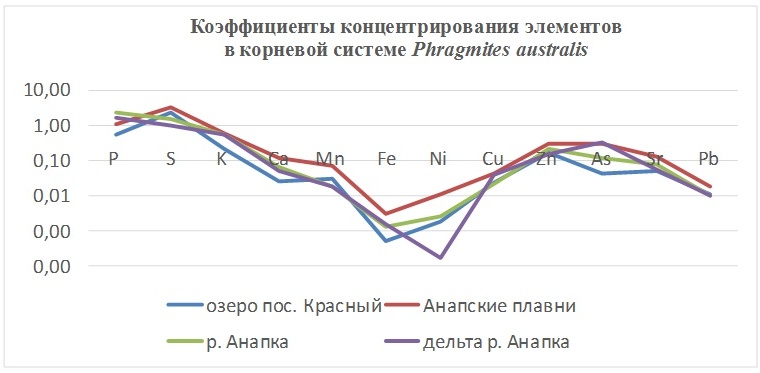
Рис. 9. Коэффициенты концентрирования элементов в корневой системе Phragmites australis.
Таблица 13. Коэффициенты биологического поглощения элементов
отмершая наддонная часть Phragmites australis /вода
(Анапский район, 2013 г., n=18,
метод определения элементов – рентгенофлуоресцентный анализ)
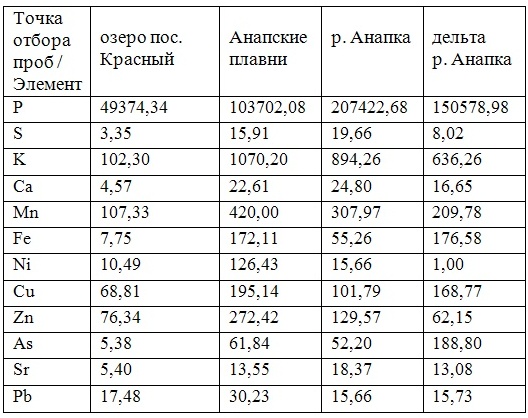
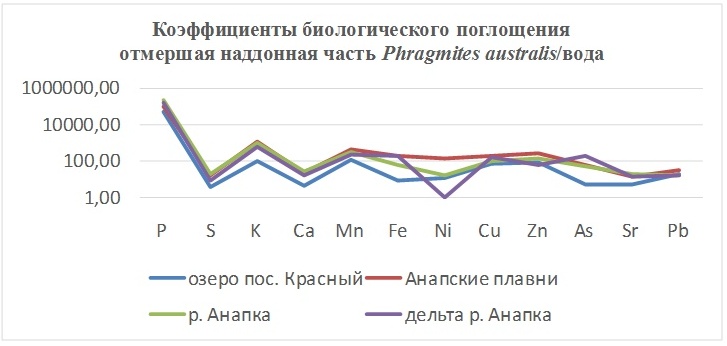
Рис. 10. Коэффициенты биологического поглощения элементов отмершая наддонная часть
Phragmites australis / вода.
Таблица 14. Коэффициенты биологического поглощения элементов
живая наддонная часть Phragmites australis /вода
(Анапский район, 2013 г., n=18,
метод определения элементов – рентгенофлуоресцентный анализ)
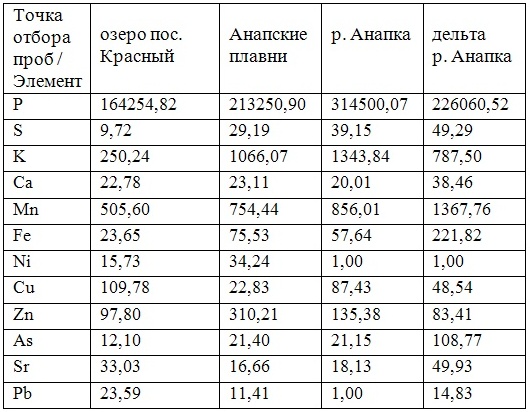
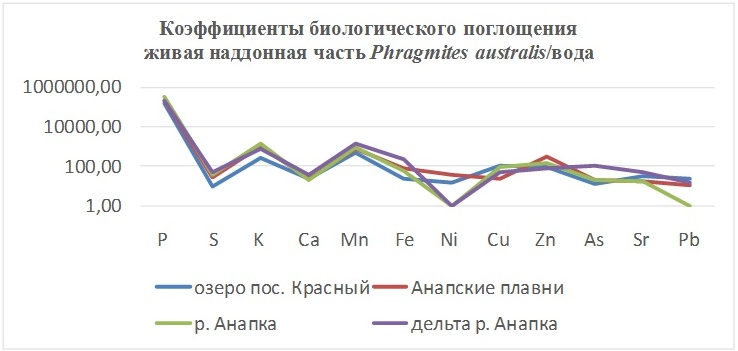
Рис. 11. Коэффициенты биологического поглощения элементов живая наддонная часть
Phragmites australis / вода.
Таблица 15. Коэффициенты биологического поглощения элементов
корневая система Phragmites australis /донные отложения
(Анапский район, 2013 г., n=18,
метод определения элементов – рентгенофлуоресцентный анализ)
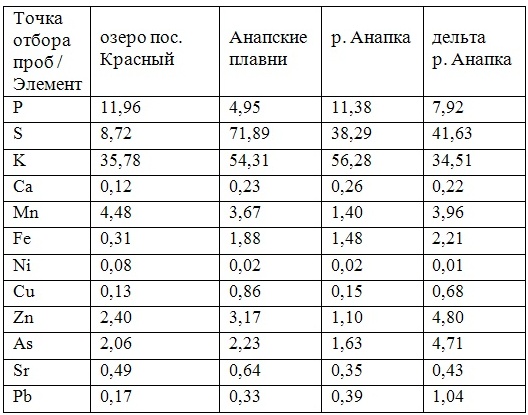
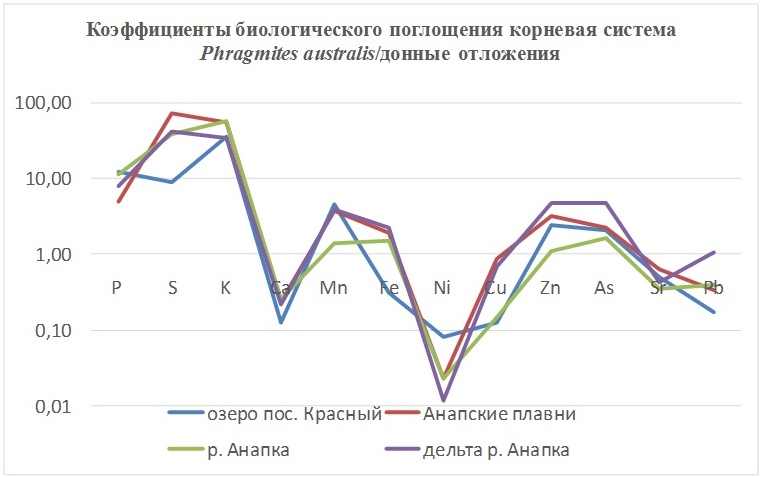
Рис. 12. Коэффициенты биологического поглощения элементов корневая система
Phragmites australis / донные отложения
Анализ графиков, отображающих изменение коэффициентов концентрирования и поглощения у Phragmites australis показывает, что все графики достаточно однотипны. Как и у морских макроводорослей, исследованных нами в анапском регионе, у тростника также наблюдается концентрирование серы и, в какой-то степени, мышьяка, что наиболее наглядно представлено на рис.8, где в живых листьях наблюдается пик. В корнях также наблюдается явно выраженный пик (рис.9), но абсолютные значения этого коэффициента не доходят до единицы. Эти данные ещё раз подтверждают наши предположения о загрязнении анапского региона соединениями серы. Вероятно, это может быть связано с многолетним использованием в сельскохозяйственной практике данного региона препаратов содержащих S, в частности, при обработке виноградников.
Коэффициенты биологического поглощения также имеют достаточно однотипный характер. В листьях наблюдаются пики коэффициентов поглощения P, K, Mn, Cu, Zn. В многолетних корнях, поглощающих из донных отложений, добавляется S. Важно отметить, что ярко выраженных особенностей в графическом отображении изменений коэффициентов концентрирования и поглощения в зависимости от места сбора объектов не наблюдается.
Представленные нами данные наглядно демонстрируют возможности данного метода, позволяющего быстро определить состояние исследуемого региона. Более того, корректно и интегративно оценить содержание макро- и микроэлементов в почвах, донных отложениях и водной среде, что в дальнейшем позволит правильно эксплуатировать или реанимировать определённый регион.
Оценка минерального состава макрофитов, собранных вдоль водных объектов по разрезу от источника загрязнения до моря
Очень показательным в биогеохимических исследованиях, на наш взгляд, является оценка минерального состава водных и прибрежно-водных макрофитов, растущих вдоль водных объектов (озеро, плавни, река), расположенных вдоль некой трансекты (разреза) от потенциальных источников загрязнения до впадения этих водных объектов в море. Для этих целей в качестве основных объектов – мониторов нами были выбраны следующие макрофиты:
1. Phragmites australis (Cav.) Trin. Ex Steud – прибрежно-водное высшее растение. Гелофит, обитающий вдоль береговой зоны рек и морей. Как уже было описано выше, корневая система этого макрофита составляет более 80% биомассы всего растения. Поглощение минеральных элементов из почвы, донных отложений и воды осуществляется основными и придаточными корнями. Это растение может выдерживать экстремальные условия окружающей среды, в том числе наличие токсичных загрязняющих веществ, таких как тяжёлые металлы (Bonanno et al., 2010, Baldantoni, 2009).
2. Carex conescens L. – прибрежно-водное растение. Гигрофит, растущий по берегам рек. В отличие от Phragmites australis, это растение имеет небольшую корневую систему. В нашем случае Carex conescens поглощал минеральные элементы только из почвы.
3. Cladophora sericea (Hudson) Kutzing – мелкая неприкреплённая (нитчатая) зелёная водоросль, обитающая в прибрежной зоне песчаных пляжей. Минеральные элементы поглощаются всей поверхностью таллома. Корневая система отсутствует.
С помощью нейтронного активационного анализа было определено содержание 41 макро- и микроэлементa (Na, Mg, Al, Cl, K, Ca, Sc, Ti, V, Cr, Mn, Fe, Co, Ni, Zn, As, Se, Br, Rb, Sr, Zr, Nb, Mo, Sb, I, Cs, Ba, La, Ce, Nd, Sm, Eu, Tb, Dy, Yb, Hf, Ta, W, Au, Th и U) в образцах растительности, почвы и донных отложений, отобранных на 7 станциях вдоль разреза «Антропогенный источник загрязнения – Анапский городской пляж» (Kamnev et al., 2014)
В прибрежной зоне нами был выбран модельный полигон, включающий городскую свалку на вершине холма Красный и все виды водоёмов (озеро, река, тростниковые поймы), локализованные ниже склона холма (под свалкой). Эти водные объекты образуют неделимый водосбор – типичную малую реку Анапка, которая впадает в море в зоне отдыха города (центральный городской пляж). Образцы макрофитов (зеленой водоросли Cladophora sericea, живые, отмершие листья и корни Phragmites australis и Carex conescens) (n=18), соответствующие образцы почв (n=16) и донных отложений (n=9) были отобраны на 7 станциях, расположенных по разрезу недалеко от г. Анапа летом 2013 года. Места отбора проб приведены на рис. 13.
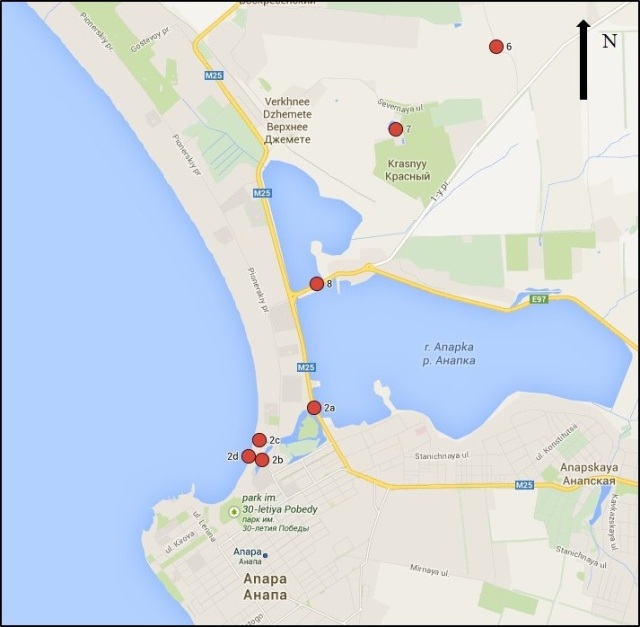
Рис.13. Точки отбора проб – трансекта в прибрежной зоне г. Анапа.
При расположении элементов в порядке убывания концентраций в изученных образцах, можно наблюдать изменение тенденции расположения элементов в рядах убывания в зависимости от типа образца:
в почве: Ca > Al > Fe, S> K > Na > Mg > Ti > Cl > Mn, Sr > Ba > Zn, Zr > V > Cr, Rb, Br > Ce > Ni > I, As, Nd > La > Co > Sc, Th, Dy, Mo, Nb, Hf > Sm, Cs, U > Yb, W, Sb > Eu > Tb, Ta, Se > Tm > Au;
в донных отложениях: Ca > Al > Fe, S> K > Na > Mg > Ti > Cl > Sr > Mn > Ba > Zr > Zn >V,Cr > Rb > Br, Ce > Ni > I, As, La > Nd > Co > Sc, Th, Mo > Dy > Hf, Nb, Sm, Cs, U > Yb, W, Sb > Eu, Tb, Ta, Se > Tm> Au;
в Phragmites australis: K > Cl > S> Na, Ca > Fe > Mg, Al > Mn > Ti > Sr, Br > I, Zn > Ba> Rb > As, Cr, Ce > V, Ni > Co, Mo > Nb, La, W > U, Yb, Sc > Th, Tm, Hf > Cs, Sm > Eu >Tb, Ta > Au;
в Carex conescens: Ca > Cl > Na > K > S > Fe > Al > Ti, Sr, Mg > Br, Zn, Ba > I > Cr > Rb, V > As, Ni > Co, Ce > Mo, Nb, La > Yb > U, W, Sc > Th, Tm > Hf, Cs, Sm, Eu > Tb, Ta > Au;
в Cladophora seriace: Ca > K > Cl > S, Al > Na, Fe > Mg > Sr > Ti, Br > Mn > Ba, I > Zn > Rb, V, As, Ce > Ni, Cr, La > Co > Sc > Th > Nb, Mo, Hf, W > Cs, Sm > U, Yb, Sb, Se > Eu, Tb, Tm, Ta > Au.
Пространственная изменчивость содержания макро- и микроэлементов в образцах макрофитов, почвы и донных отложений.
Макрофиты
В живых листьях Phragmites australis максимальные концентрации S, Cl, K, Ti, Br, As, Rb, Nb наблюдались на станции 2b, высокие значения Na, Ca, Cr, Fe, Ba, REE, W – на станции 2с. Концентрации Mg, Al, V, Mn, Se, Sr, Mo, I были в 2-5 раза выше на станции 7 по сравнению с другими точками отбора проб.
В корнях Phragmites australis максимальные концентрации большинства элементов (за исключением Cl, K и Cr) наблюдались на станции 7. Кроме того, концентрации таких элементов, как Al, Cs, V, Mn, Fe, Co, Mo, Sb, Cs, La, Sm, Th и U на этой станции достигали значений, в 30 и более раз превышающих аналогичные показатели для других точек отбора проб. Наиболее высокие концентрации K, Cr и Cl обнаружены в корнях Phragmites australis на станциях 2b и 2с.
Концентрации большинства элементов (за исключением Ca, Mn and Br) в зелёных морских водорослях Cladophora seriace со станций 2c и 2d были очень похожи. Самые высокие значения Ca, Mn and Br были обнаружены в Cladophora seriace со станций 2c и 2d.
Донные отложения
Максимальные концентрации большинства минеральных элементов (за исключением Br и Sr) наблюдались в донных отложениях со станций 7 и 8 (Рис. 16). Самые высокие концентрации Br и Sr были обнаружены на станции 2b.
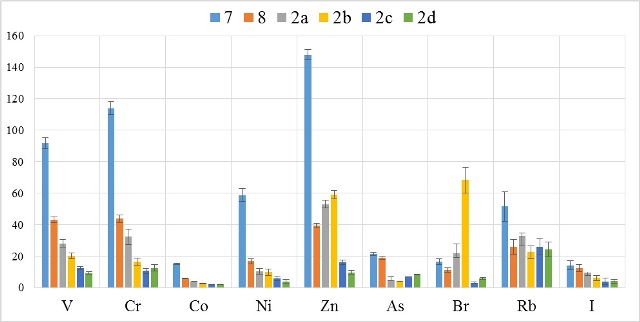
Рис. 14. Пространственное изменение некоторых минеральных элементов (мг/ кг) в донных отложениях (станции 7, 8, 2а, 2b, 2c, 2d).
Почвы
Максимальные концентрации большинства минеральных элементов (за исключением Br) наблюдались в почвах на станции 6 и 7 (рис. 17). Интересно отметить, что наибольшая концентрация Zn была обнаружена на станции 6 (где находится городская свалка).
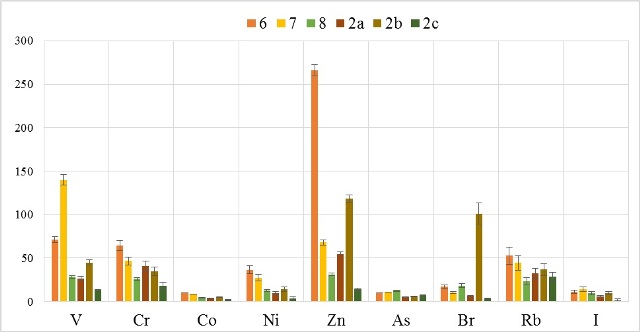
Рис. 15. Пространственное изменение некоторых минеральных элементов (мг/кг) в почвах (станций 6, 7, 8, 2а, 2b, 2с)
Межвидовые изменения накопления элементов
Результаты сравнения элементного накопления в живых листьях и корнях Phragmites australis и Carex conescens приведены в таблице 16.
Таблица 16. Соотношение между концентрацией минеральных элементов
в живых листьях и корнях Phragmites australis и Carex conescens.
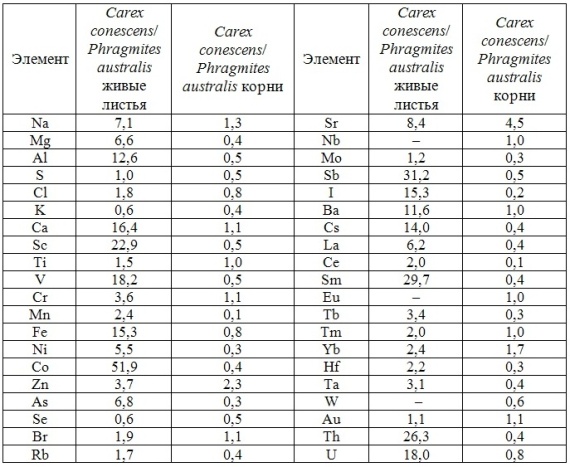
Концентрации всех элементов (за исключением S, K, Se, Au) в живых листьях были выше у Carex conescens, чем у Phragmites australis. Наибольшие различия наблюдались в накоплении Ca, V, Sc, Fe, Co, Sb, I, Sm, Th и U (более чем в 15 раз). Обратная закономерность была обнаружена для накопления элементов в корнях, т.е. концентрации одних и тех же элементов в корнях тростника обычно были выше, чем в корнях осоки. Исключение составили Na, Zn, Sr и Yb.
Важно отметить, что концентрации Al, Ca, Sc, V, As, Br, Sr, La, Ce, Sm, Th в Cladophora sericea в 10 и более раз выше по сравнению с их содержанием в Phragmites australis и Carex conescens.
Накопление элементов различными органами Phragmites australis
Различия накопления элементов в зависимости от органа растения (живые и отмершие листья, а также корни) было изучено для Phragmites australis, собранных на станции 7. В результате исследований были получены следующие закономерности: для Na, Mg, Al, Sc, V, Fe, Ni, Co, As, Nb, Mo, Sb, I, Ba, Cs, La, Ce, Sm, Eu, Tb, Tm, Ta, Au, Th, U максимальные концентрации наблюдались в корнях, а минимальные в живых листьях; для S, Ca, Ti, Mn, Zn, Se, Br, Rb, Hf – корни > живые листья > отмершие листья; для Cr, Yb, W, Sr – отмершие листья > живые листья > корни. Максимальные концентрации Cl и K были обнаружены в живых листьях, минимальные – в отмерших листьях и корнях.
Эти исследования показали, что элементное распределение среди различных образцов уменьшалось в следующем порядке: почвы, донные отложения> корни> листья. Имеются данные, что укорененные макрофиты, такие как Phragmites australis, поглощают больше элементов из донных отложений, чем из воды (Baldantoni и др., 2009; Bonanno et al., 2010; Bonanno, 2011). Полученные результаты подтвердили, что Phragmites australis является преимущественно корневым биоаккумулятором. С другой стороны, было замечено, что большинство элементов в Carex conescens было накоплено в листьях.
Самые высокие концентрации большинства минеральных элементов были обнаружены в образцах со станции 7. Эта станция расположена на расстоянии 1 км от антропогенного источника загрязнения, который является источником загрязнения атмосферного воздуха, почвы, поверхностных и грунтовых вод. Поверхностный сток проникает в местную речную систему реки Анапка, доставляя загрязняющие вещества к побережью и осаждая их на пути. Таким образом, почва, донные отложения и макрофиты, расположенные вдоль русла реки, работают естественными фильтрами, уменьшая концентрации элементов. Аналогичные закономерности были выявлены другими авторами (Камбарова, 2011).
Для более глубокого понимания процессов, происходящих с минеральными элементами в растительных объектах и средах их обитания, нами были проведены сравнения с литературными данными (табл. 17). В результате этого анализа нами не было выявлено чётких общих закономерностей. Для каждого региона были выявлены свои закономерности, связанные с биогеохимической обстановкой местности. Так, например, концентрации большинства минеральных элементов были выше в листьях и корнях Phragmites australis, отобранных в горном озере в Италии (Baldantoni et al., 2009) и в устье самой длинной сицилийской реки (Bonanno et al., 2010; Bonanno, 2011) по сравнению с нашими данными. Исключениями были концентрации Ti, Mn, As, Sb и Ti, V, As, Se, значения которых были выше в корнях и листьях, соответственно. Концентрации Co, Rb, Zn и Th были выше в Carex conescens, отобранной в Германии в ботаническом саду (Horovitz et al., 1974) по сравнению с нашими результатами; обратная тенденция наблюдалась для Fe, Cr, Cs и Sc. Концентрации Mg, Mn, Na и Zn были выше в Cladophora sericea из Балтийского моря (Bojanowski, 1973); концентрации Ca, Fe, Ni, Co и Sr были ниже, чем в настоящем исследовании. Концентрации всех элементов (за исключением Mn и V) опубликованные Дьяченко с коллегами (Dyachenko at al., 2014) для почв Юга России были выше, чем в образцах почв, собранных нами с наиболее загрязнённой станции около антропогенного источника загрязнения.
Таблица 17. Литературные данные о концентрации минеральных элементов (мг/кг)
в Phragmites australis, Carex conescens, Cladophora seriace и почвах.
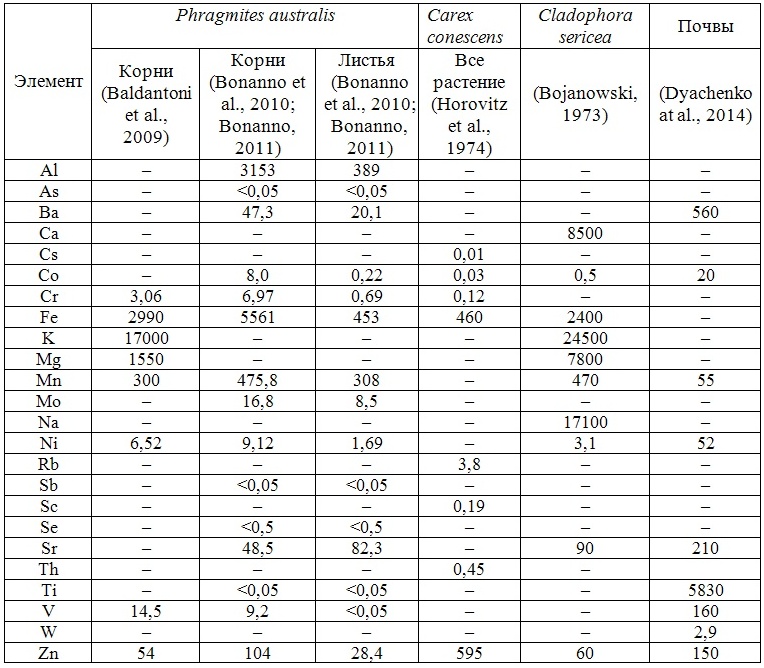
Коэффициенты корреляции концентрации минеральных элементов в различных органах Phragmites australis и в почве показали положительную корреляцию Mg, Al, V, Mo, I между живыми листьями, корнями и почвой (табл. 18). Противоположенные коэффициенты корреляции были найдены для Na, Fe, Br, U в живых и отмерших листьях. Не исключено, что это может быть связано с высокой степенью физиологического регулирования аккумулирования этих элементов в живых органах растений.
Таблица 18. Корреляция между средними концентрациями некоторых минеральных элементов
в различных органах Phragmites australis и концентрацией в почве.
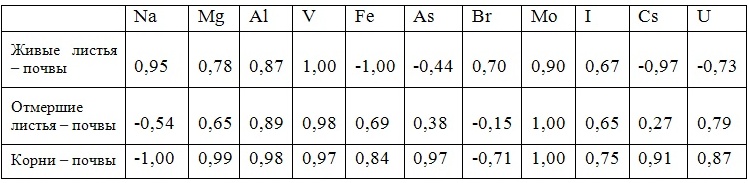
Аналогичные коэффициенты корреляции наблюдались между концентрациями минеральных элементов в различных органах Phragmites australis и в донных отложениях (табл. 19). Сильная положительная корреляция была найдена между корнями и донными отложениями (за исключением Na, Cl and Br).
Таблица 19. Корреляция между средними концентрациями некоторых минеральных элементов
в различных органах Phragmites australis и концентрацией в донных отложениях.

Таким образом, представленные нами данные могут служить наглядным примером целесообразности выбора водных макрофитов для решения многих биогеохимических задач. Кроме того, эти данные хорошо демонстрируют экологическое и видовое разнообразие физиологических механизмов накопления минеральных элементов из окружающей среды.
Важно отметить, что нами впервые (для стратегически важного региона г. Анапа) были определены концентрации Na, Mg, Al, Cl, K, Ca, Sc, Ti, V, Cr, Mn, Fe, Co, Ni, Zn, As, Se, Br, Rb, Sr, Zr, Nb, Mo, Sb, I, Cs, Ba, La, Ce, Nd, Sm, Eu, Tb, Dy, Yb, Hf, Ta, W, Au, Th и U в различных образцах: живые и отмершие листья и корни Phragmites australis и Carex conescens, Cladophora seriace и соответствующих почвах и донных отложениях, собранных вдоль разреза от места потенциального загрязнения (городская свалка) до моря, находящегося в 4,5 км от него. Эти данные также могут служить примером о возможном использование макрофитов для задач такого рода. Например, из полученных нами данных видно, что самые высокие концентрации большинства элементов были найдены в корнях растений, почвах и донных отложениях со станции 7, находящейся в 1 км от антропогенного источника загрязнения как почвы, так и поверхностных и грунтовых вод. Исследования показали, что распределение элементов среди различных образцов уменьшалось в следующем порядке: почвы, донные отложения > корни Phragmites australis> живые листья Phragmites australis. С другой стороны, было замечено, что большинство элементов в Carex conescens было накоплено в листьях.
Концентрации всех элементов (за исключением S, K, Se, Au) были выше в живых листьях у Carex conescens, чем у Phragmites australis. Самые большие различия наблюдались в накоплении Ca, V, Sc, Fe, Co, Sb, I, Sm, Th и U (более чем в 15 раз). Обратная закономерность была найдена в накоплении элементов в корнях Carex conescens и Phragmites australis. Концентрация в корнях Phragmites australis обычно были выше (за исключением Na, Zn, Sr и Yb), чем в корнях Carex conescens.
Сильные положительные корреляции между концентрациями элемента в Phragmites australis и окружающей среде предполагают, что этот вид макрофита потенциально полезен для контроля загрязнения в целом и большинства исследованных минеральных элементов в частности.
Не менее удобными организмами – мониторами могут служить и морские макрофиты, например, представители рода Cystoseira, Cladophora и Zostera, которые достаточно чётко отражают не только состояние биогеохимического состояния водной среды, но и региона в целом.
Следовательно, минеральный состав макрофитов может быть интегративно использован в качестве важнейшего показателя оценки биогеохимического состояния окружающей среды и их вклада в биогеохимический обмен Мирового океана.
Таким образом, настоящая работа показывает, что идеи Дмитрия Анатольевича Сабинина были актуальны не только в середине прошлого столетия, но актуальны и в настоящее время, а макрофиты (в т.ч. цистозейра, которую он выбрал в качестве объекта исследований), являются действительно удобными модельными объектами для решения различных биогеохимических задач.
Список литературы
1. Базилевич Н.И. Биологическая продуктивность экосистем Северной Евразии. – М.: Наука, 1993. — 293 с.
2. Базилевич Н.И., Гребенщиков О.С., Тишков А.А. Географические закономерности структуры и функционирования экосистем. – М.: Наука, 1986. – 298 с.
3. Базилевич Н.И., Титлянова А.А., Смирнов В.В. Методы изучения биологического круговорота в разных природных зонах. – М.: Мысль, 1978. – 182 с.
4. Башкин В.Н. Биогеохимия. М. – М.: Научный мир, 2004 . – 582 с.
5. Богатырев Л.Г. Геохимические спектры как одна из форм анализа состояния объектов окружающей среды // Профессиональный лекторий Всероссийский молодежный научный форум «Шаг в будущее». Симпозиум 2. Естественные науки и современный мир. — РОО НТА АПФН, Москва, 2008. – С. 36-39.
6. Богатырев Л.Г. Основные концепции, законы и принципы современного почвоведения. – М.: Макс Пресс, 2015. – С. 196.
7. Богатырев Л.Г., Рыжова И.М. Биологический круговорот и его роль в почвообразовании. – М.: Изд-во Московского университета, 1994. – 80 с.
8. Богатырев Л.Г., Цветнова О.Б., Щеглов А.И. Биогеохимия – от биологического круговорота к биогеохимическим циклам // Проблемы биогеохимии и геохимической экологии. – 2007. – Т. 2, № 4. – 2007 – С. 5-15.
9. Богатырев Л.Г., Макаров О.А., Семенюк О.В., Матышак Г.В. О некоторых тенденциях в изучении биосферы // Экология. – 2004. – № 1. – С.3-12.
10. Богатырев Л.Г., Телеснина В.М. Словарь терминов и показателей, используемых при изучении биологического круговорота. – М.:МАКС Пресс, 2010. – 182 с.
11. Бурдин К. С. Основы биологического мониторинга. – М., МГУ, 1985. – 156 с.
12. Бяллович Ю.П. Биогеоценологические основания теории систем лесов // Проблемы биогеоценологии. – М.: Наука, 1973. – С. 47–58.
13. Бяллович Ю.П. Системы биогеоценозов // Проблемы биогеоценологии. – М.: Наука, 1973. – С.37–46
14. Вернадский В.И. Живое вещество в химии моря. – Пг: Науч. хим.-техн. изд-во, 1923. – 36 с.
15. Вернадский В.И. Проблемы биогеохимии. 1. Значение биогеохимии для изучения биосферы. – Л.: Изд-во АН СССР, 1934. – 47 с.
16. Вернадский В.И. Биогеохимические очерки. – М.; Л.: Изд-во АН СССР, 1940. – 241 с.
17. Вернадский В.И. Проблемы биогеохимии. Вып. 4. О правизне и левизне. – М.; Л.: Изд-во АН СССР, 1940 – 16с.
18. Вернадский В.И. Избранные соч. Т.4, кн. 2. История природных вод. – М., Изд-во АН СССР, 1960. – 657 с.
19. Вернадский В.И. Проблемы биогеохимии. Труды биогеохимической лаборатории. – М., Наука, 1980. – 320 с.
20. Вершинин А.О. Жизнь Чёрного моря. – Краснодар: Когорта, 2007. – 191 с.
21. Виноградов А.П. Средние содержания химических элементов в главных типах изверженных горных пород земной коры // Геохимия. – 1962. – № 7. – С. 555-571.
22. Возжинская В.Б., Камнев А.Н. Эколого-биологические основы культивирования и использования морских донных водорослей. – М.: Наука, 1994. – 202 с.
23. Генкель П.А. Дмитрий Анатольевич Сабинин, 1889-1951 – М.: Наука, 1980. – 183 с.
24. Гильманов Т.Г., Базилевич Н.И. Концептуальная балансовая модель круговорота органического вещества в экосистеме как теоретическая база мониторинга // Теоретические основы и опыт экологического мониторинга. – М.: Наука, 1983. – С. 7-23.
25. Глазовская М.А. Геохимические основы типологии и методики исследования природных ландшафтов. – М., Изд-во Моск. ун-та. 1964. – 230 с.
26. Глазовская М.А. Методологические основы оценки эколого-геохимической устойчивости почв к техногенным воздействиям. – М., Изд-во Моск. ун-та. 1997. – 102 с.
27. Дедю И.И. Экологический энциклопедический словарь. – Кишинев. – 1989. – 406 с.
28. Добровольский В.В. Основы биогеохимии. – М.: Издательский центр Академия, 2003 г. – 398 с.
29. Зайцев Ю.Т. Введение в экологию Черного моря. – Одесса: Эвен, 2006. – 224 с.
30. Зинова А.Д. Определитель зелёных, бурых и красных водорослей южных морей СССР. – М.: Наука, 1967. – 393 с.
31. Казначеев В.П. Учение о биосфере (Этюды о научном творчестве В. И. Вернадского). – М.: Знание, 1985 – 80 с.
32. Калабухов Н.И. Сохранение энергетического баланса организма как основы процесса адаптации // Журн. общей биол. 1946, 7, вып. 6. С.417-434.
33. Калугина-Гутник А.А. Фитобентос Черного моря. – Киев: Наукова Думка,1975. – 247 с.
34. Камбарова Е.А. Изучение проблем устойчивого развития рекреационных центров Краснодарского края // Материалы IV всероссийской научно-практической конференции «Курортно-рекреационный комплекс в системе регионального развития: инновационные подходы», Краснодар, 15 апреля 2011 г. – Краснодар: КубГУ, 2011. – P. 333.
35. Камнев А.Н. Структура и функции бурых водорослей. – М.: Изд. МГУ, 1989. – 200 с.
36. Камнев А.Н., Жигалова Т.В., Бассарская Е.М., Аверчева О.В. Экологическая физиология водных фототрофных организмов. Фотосинтез. – М.: Издательство Московского университета, 2013. –134 с.
37. Камнев А.Н., Мессинева Е.М., Рудченко М.Н., Силкин В.А. Возрастные изменения морфофункциональных характеристик представителей рода бурых водорослей Cystoseira Черного и Японского морей // Вопросы современной альгологии. – 2012. – № 2 (2). URL: http://algology.ru/148
38. Ковалевский А.Л. О биогеохимических параметрах растений и некоторых особенностях изучения их. // Биогеохимия растений. – Улан-Удэ: Бурят. Книжное изд-во, 1969. – С. 195-214.
39. Ковалевский А.Л. Биогеохимия растений. – Новосибирск: Наука, 1991. – 293 с.
40. Ковальский В.В. Геохимическая экология. – М.: Наука, 1974. – 274 с.
41. Ковальский В.В. Геохимическая экология – основа системы биогеохимического районирования // Труды биогеохим. лабор. № 17. – М.: Наука, 1978. – С. 3-20.
42. Ковальский В.В. 60 лет биогеохимии. // Труды БГХ лаборатории, № 20. – М.: Наука, 1985. – С. 5–20.
43. Ковда В.А. Основы учения о почвах. Общая теория почвообразовательного процесса (в 2-х кн.) – М.: Наука, 1973. – 448 с. + 468 с.
44. Ковда В.А. Биогеохимия почвенного покрова. – М. Наука, 1985. – 263 с.
45. Коновалов С.К., Рябинин А.И. Водородный показатель (рН) вод Чёрного моря // Метеорология и гидрология. – 1987. – № 10. – С. 75-81.
46. Колосов И.И. , Трубецкова О. М. // В кн: Д.А. Сабинин. Физиологические основы питания растений. – М.: Изд. АН СССР,1955. – С. 486.
47. Крапивин В.Ф., Свирежев Ю. М., Тарко А.М. Математическое моделирование глобальных биосферных процессов. – М.: Наука, 1982. – 271 с.
48. Лукина Л.Ф. , Смирнова Н.Н. Физиология высших водных растений. – Киев: Наукова Думка, 1988. – 186 с.
49. Максимова Ю.Г., Русакова Е.С., Толпешта И.И., Соколова Т.А., Близнецов К.Ю., Мокроусова П.А. Специфика кислотно-основной буферности почв в ризосфере ели и в горизонте АЕ подзолистой почвы // Вестник Московского Университета. – 2012. – № 2. – С. 28-34.
50. Одум Ю. Экология: В 2-х т. – М.: Мир, 1986. – Т. 1. – 328 с., Т. 2. – 376 с.
51. Орлов Д.С., Бирюкова О.Н., Суханова Н.И. Органическое вещество почв Российской Федерации. – М.: Наука, 1996. – 258 с.
52. Перельман А.И. Биокосные системы Земли. – М.: Наука, 1977. – 163 с.
53. Перельман А.И. Геохимия. – М.: Высшая школа, 1989. – 531 с.
54. Перельман А.И., Касимов Н.С. Геохимия ландшафта. – М.: Астрея-2000, 1999. – 764 с.
55. Плотников Г.К. Чёрное море. – Краснодар: Кубанское книжное издательство, 2007. – 104 с.
56. Полынов Б.Б. Избранные труды. – М.: Изд-во АН СССР, 1956. – 751 с.
57. Распопов И.М. Высшая водная растительность больших озёр Северо-Запада СССР. – Л., 1985. – 199 с.
58. Реймерс Н. Ф. Экология. Теории, законы, правила, принципы и гипотезы. – М.: Россия молодая, 1994. – 366с.
59. Рьюз М. Философия биологии – М: Прогресс, 1977. – 320 с.
60. Сабинин Д.А. Физиологические основы питания растений. – М.: Изд-во АН СССР, 1955. – С. 512.
61. Сабинин Д. А., Щапова Т. Ф. Темп роста, возраст и продукция Cystoseira barbata в Черном море // Тр. Ин-та Океанолог. – 1954. – Т. 8. – С. 119–146.
62. Снакин В.В. Биогенный круговорот химических элементов и подходы к его изучению // Биогеохимический круговорот веществ биосфере. – М.: Москва, 1987. – С. 50-56.
63. Соколов И.А., Конюшков Д. Е. О законах генезиса и географии почв // Почвоведение. - 2002. – № 7. – С. 777-788.
64. Сукачев В.Н. Избранные труды. В 3 т. – Т. 1. Основы лесной типологии и биогеоценологии. – Л.: Наука, 1972. – 420 с.
65. Таргульян В.О. Почвообразование и выветривание в холодных гумидных областях. – М.: Наука, 1971. – 270 с.
66. Тиссен С. Геохимические и фитологические связи в свете в свете прикладной геофизики. // В кн.: Геохимические методы поисков рудных месторождений. – М., 1954. – С. 325-372.
67. Титлянова А.А. Системное описание круговорота веществ. Основные понятия и количественные параметры // Экология. – 1984. – №1. – С. 58-59.
68. Титлянова А.А., Ляпунов А.А. Системный подход к изучению обменных процессов в биогеоценозе // Ботанический журнал. – 1974. – Т. 59, № 8. – С.1081-1092.
69. Титлянова А.А. Процессы круговорота углерода и азота в агроценозах // Агроценозы степной зоны. – Новосибирск: Наука, 1984. – 143 с.
70. Тюрюканов А.Н. Избранные труды. – М.: РЭФИА, 2001. – С.213-218.
71. Федченко Б.А. Высшие растения // Жизнь пресных вод. – М.; Л. Изд-во АН СССР,1949. – Т. 2. – С. 311-338.
72. Ферсман Ф.Е. Геохимия. – Т. I –IV. – М.: Изд-во АН СССР, 1955, 1958, 1959.
73. Фокин А.Д., Черникова И.Л., Ибрагимов К.Ш., Сюняев Х.Х. Роль растительных остатков в обеспечении растений зольными элементами на подзолистых почвах. // Почвоведение. 1979. № 6. С. 53-61.
74. Фокин А.Д. Почва, биосфера и жизнь на Земле. – М., Наука, 1986. – 176 с.
75. Шенников А. П. Экология растений. – М., 1950. – 375 с.
76. Шилов И.А. Экология. – М.: Высш. школа. – 2001. – 512 с.
77. Baldantoni, D., Ligrone, R., Alfani, A. Macro-and trace-element concentrations in leaves and roots of Phragmites australis in a volcanic lake in Southern Italy. // Journal of Geochemical Exploration. – 2009. – Vol. 101(2). – P. 166-174.
78. Bonanno, G. Trace element accumulation and distribution in the organs of Phragmites australis (common reed) and biomonitoring applications. // Ecotoxicology and environmental safety. – 2011. – Vol. 74(4). – P. 1057-1064.
79. Bonanno, G., Lo Giudice, R. Heavy metal bioaccumulation by the organs of Phragmites australis (common reed) and their potential use as contamination indicators. // Ecological Indicators. – 2010. – Vol. 10(3). – P. 639-645.
80. Bojanowski R. The occurrence of major and minor chemical elements in the more common Baltic seaweed. // Oceanologia. – 1973. – 2. – P. 81-152.
81. Bunkova O., Kamnev A., Yakovlev A., Suhova T., Shahpenderyan E. Studying of mineral structure of Cystoseira barbata in a sublittoral zone of the northeast coast of the Black Sea. // World Conference on Marine Biodiversity, Qingdao, China, 12‐16 October 2014. – Р. 76.
82. Dyachenko, V., Matasova, I., & Ponomareva, O. The Trace Elements Concentrations Dynamics in the Soil Landscapes of the Southern Russia. // Universal Journal of Geoscience. – 2014. – Vol. 2(1). – P. 28-34
83. Horovitz C. T., Schock H. H., Horovitz-Kisimova L. A. The content of scandium, thorium, silver, and other trace elements in different plant species. // Plant and Soil. – 1974. – Т. 40, №. 2. – P. 397-403.
84. Kamnev A.N., Frontasyeva M.V., Kravtsova A.V., Nekhoroshkov P.S., Bunkova O.M., Stukolova I.V., Yakovlev A.S., Zinicovscaia I.I., Yushin N.S. Assessment of elemental composition of macrophytes, soil and bottom sediments along a transect in the coastal zone of Anapa, studied by neutron activation analysis // Issues of modern algology. – 2014. – № 3S (7S). Proceedings of the International Conference «Ecological physiology of aquatic phototrophic: distribution, stocks, chemical composition and use» III Sabininskie reading. Part2. URL: http://algology.ru/647
85. Milchakova N.A. Marine plants of the Black Sea. An illustrated Field Guide. – Sevastopol, 2011. – 144 p.
86. Soudzilovskaia N.A., van_der_Heijden M.G.A, Cornelissen J.H.C, Makarov M.I., Onipchenko V.G., Maslov M.N., Akhmetzhanova A.A., van_Bodegom P.M. Quantitative assessment of the differential impacts of arbuscular and ectomycorrhiza on soil carbon cycling // New Phytologist, Blackwell Publishing Inc. (United Kingdom). 208, № 1. P. 280-293. doi:10.1111/nph.13447.
87. Vershinin A., Kamnev A. Harmful algae in Russian evropean coastal waters // 9th Int. Conf. on Harmful algal blooms: Proceedings. Hobart, Australia, 7-11 February 2000. – UNESCO. 2001. – P. 112-114.
Статья поступила в редакцию 5.12.2015
D. A. Sabinin's ideas and their embodiment.
Mineral composition of macrophytes is one of the most important indicator of their contribution to a biogeochemical exchange of the World Ocean.
A.N. Kamnev, O.M. Bunkova, L.G. Bogatyrev, I.V. Stukolova, A.S. Yakovlev
The work questions the biogeochemistry concept, and its historical evolution as an independent science. This research shows the structure of modern biogeochemistry, it discusses its goals and objectives, provides the difinition of biosphere. Some parallels in history of evolution of plant physiology, soil science and biogeochemistry are considered. In the research the special attention is given to the ideas of D.A. Sabinin, who proved importance of development of biogeochemical researches in the aquatic environment, and he suggested to use water phototrophs for those researches as they are easy to examine, especially macrophytes, marine macroscopic algae. The research gives much attention to the changes of chemical composition of coastal part of the Black Sea. The sea, including a coastal zone, is considered, on the one hand, as convenient model for a biogeochemical system analyses, and on the other, as strategically important region not only for Russia, but for Europe and Asia. This work features the range change of mineral composition and accumulation capacity of soil, ground deposits and macrophytes. Also this research provides examples of change of mineral composition of coastal and water plants, that grow from the spring region to the mouth of the small river.
Keywords: biogeochemistry; biosphere; phototrophs; water and coastal plants–macrophytes; ground deposits; soil; aquatic habitat; the Black Sea; mineral composition of macrophytes; mineral composition of ground deposits; mineral composition of the soil; mineral composition of the aquatic environment.
Об авторах
Камнев Александр Николаевич - Kamnev Alexander Nikolaevich
доктор биологических наук
ФГБОУ ВПО «Московский государственный университет им.М.В.Ломоносова», Москва, Россия (Lomonosov Moscow State University, Moscow, Russia), биологический факультет, кафедра физиологии растений
dr.kamnev@mail.ru
Бунькова Ольга Михайловна - Bunkova Olga M.
аспирант ФГБОУ ВПО «Московский государственный университет им. Ломоносова», Москва, Россия (Lomonosov Moscow State University, Moscow, Russia), факультет почвоведения, кафедра оценки земельных ресурсов.
BunkovaOlga@yandex.ru
Богатырев Лев Георгиевич - Bogatyrev Lev G.
кандидат биологических наук
доцент, ФГБОУ ВПО «Московский государственный университет им.М.В.Ломоносова», Москва, Россия (Lomonosov Moscow State University, Moscow, Russia), Факультет почвоведения, Кафедра общего почвоведения
Стуколова Ирина Владимировна - Stukolova Irina V.
ФГБОУ ВПО «Московский государственный университет им.М.В.Ломоносова», Москва, Россия (Lomonosov Moscow State University, Moscow, Russia), биологический факультет, кафедра физиологии растений
iri_sk@inbox.ru
Яковлев Александр Сергеевич - Yakovlev Alexander S.
профессор, доктор биологических наук
Заведующий кафедрой ФГБОУ ВПО «Московский государственный университет им. Ломоносова», Москва, Российская федерация (Lomonosov Moscow State University, Moscow, Russia), факультет почвоведения, кафедра оценки земельных ресурсов.
yakovlev_a_s@mail.ru
Корреспондентский адрес: Россия, 119991, Москва, Ленинские горы, Московский Государственный Университет им.М.В.Ломоносова, д. 1, стр. 12, факультет почвоведения, кафедра оценки земельных ресурсов.
ССЫЛКА НА СТАТЬЮ:
Камнев А.Н., Бунькова О.М., Богатырев Л.Г., Стуколова И.В., Яковлев А.С. Идеи Д.А. Сабинина и их воплощение. Минеральный состав макрофитов как важнейший показатель их вклада в биогеохимический обмен Мирового океана // Вопросы современной альгологии. 2015. № 3 (10). URL: http://algology.ru/933
Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию и мы ее вам с удовольствием пришлем бесплатно.
Адрес - info@algology.ru
При перепечатке ссылка на сайт обязательна
К № 3 (10)
Карта сайта

|
|
|
