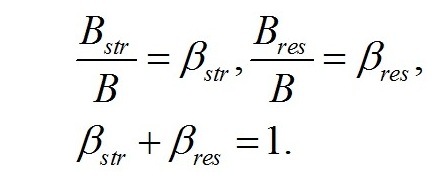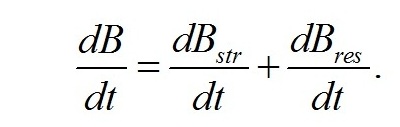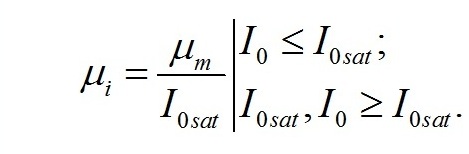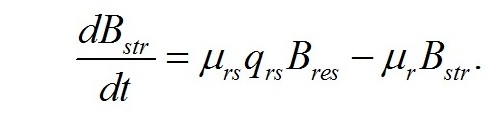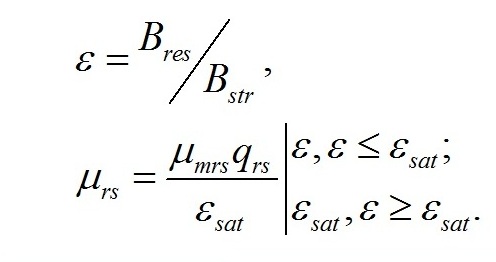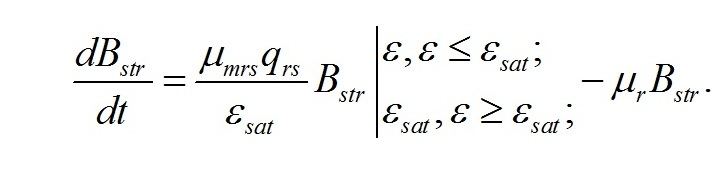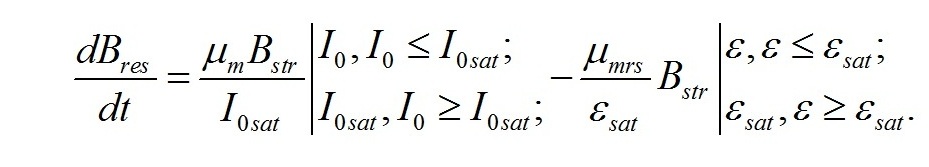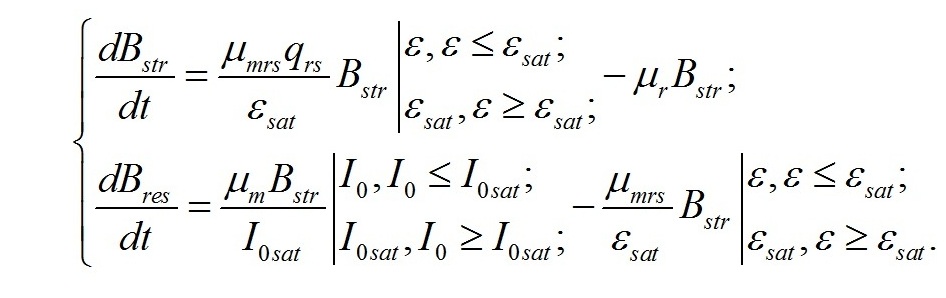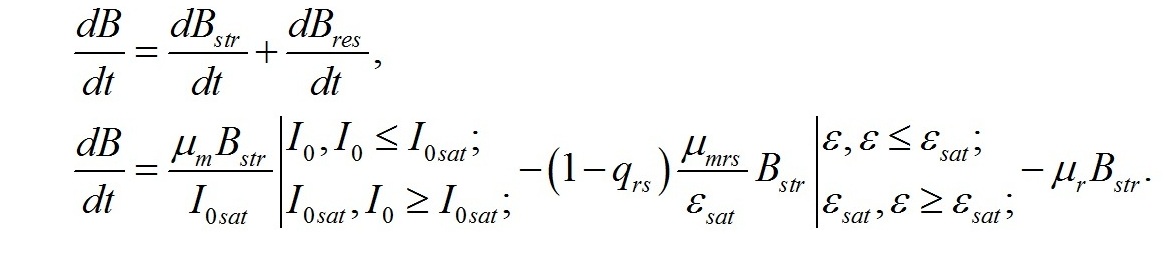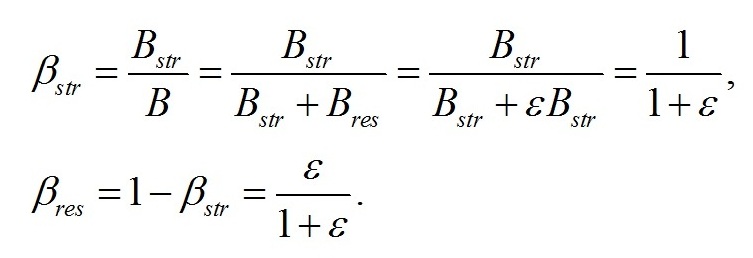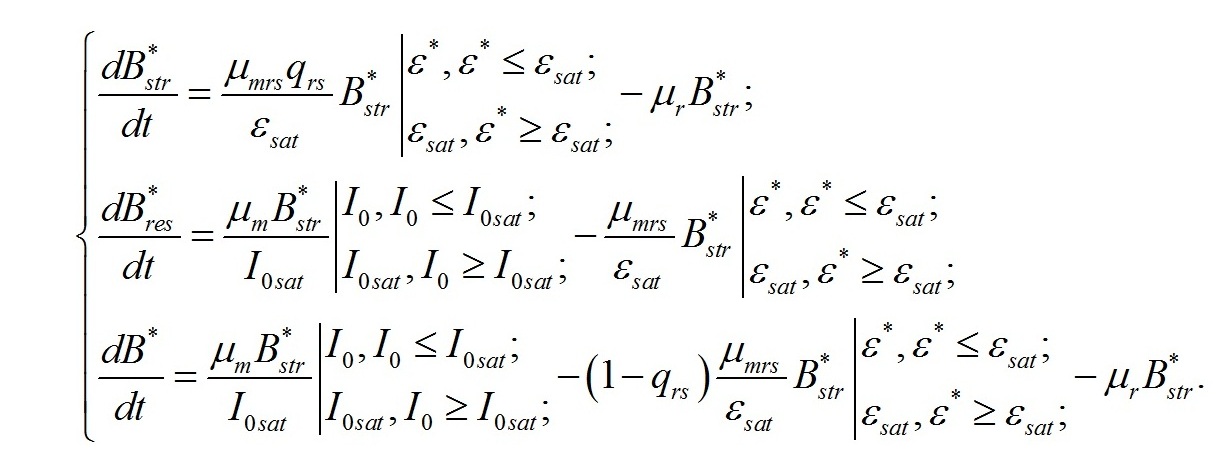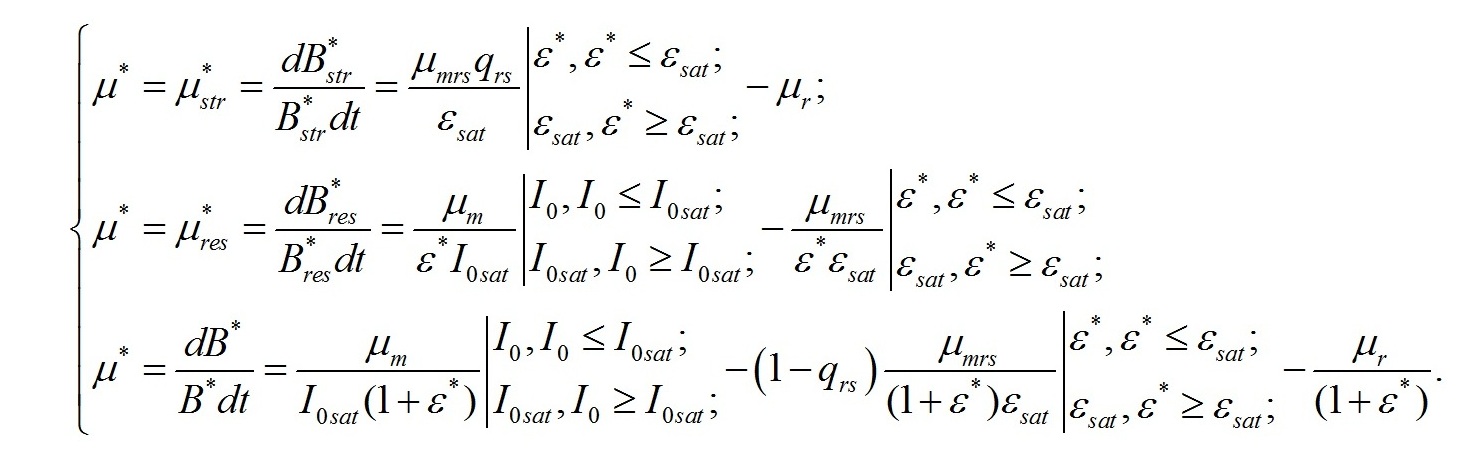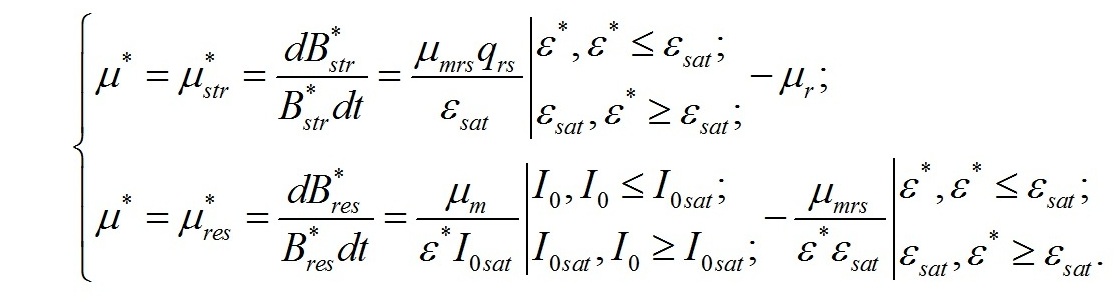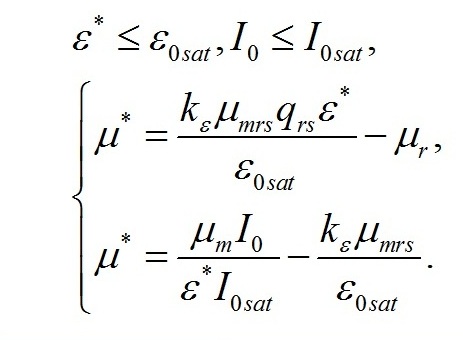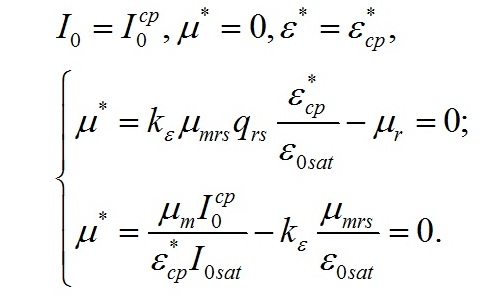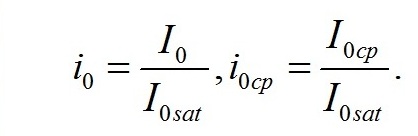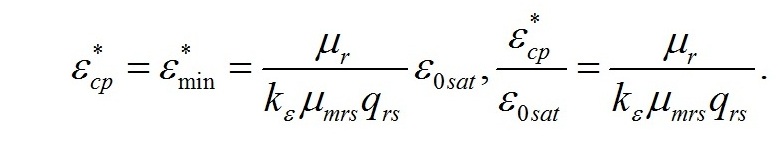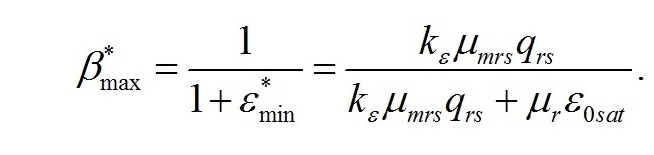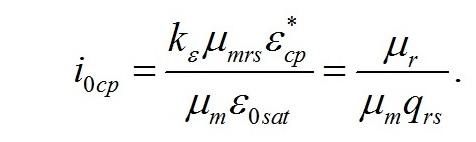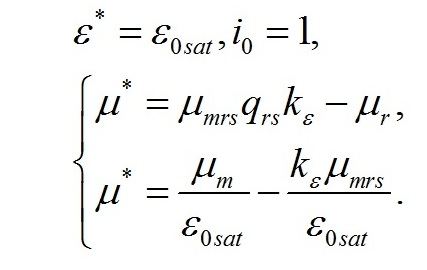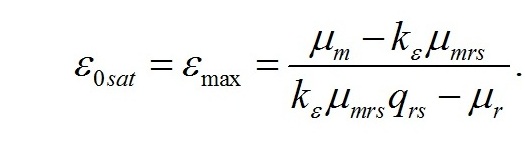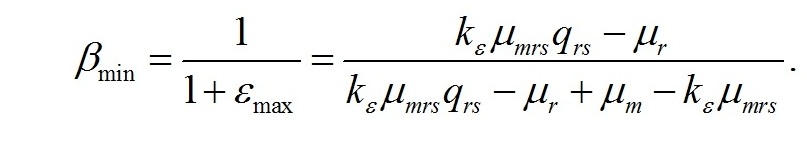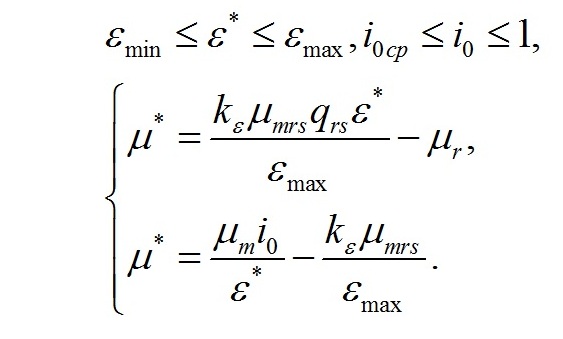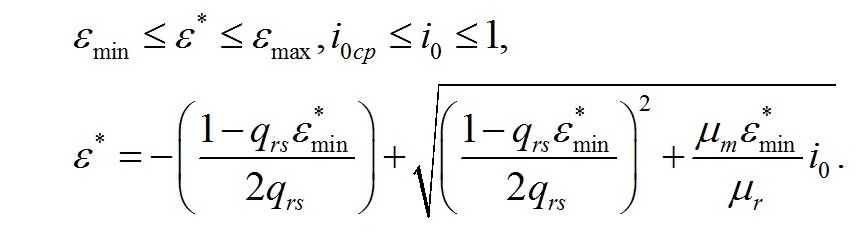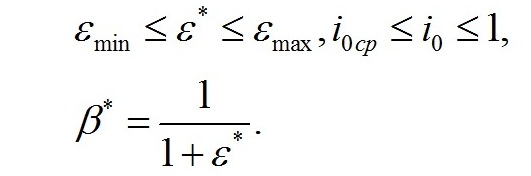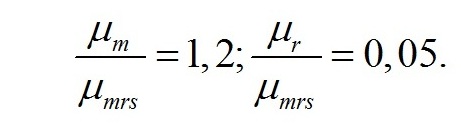|
|
|
Влияние света на макромолекулярный состав микроводорослей в непрерывной культуре невысокой плотности (Часть 1)
Тренкеншу Р.П. Rudolf P. Trenkenshu
Институт морских биологических исследований им. А.О. Ковалевского (г. Севастополь)
УДК 582.232:581.121
Показано, что в динамически равновесной (непрерывной) культуре невысокой плотности макромолекулярный состав биомассы микроводорослей существенно зависит поверхностной освещенности культуры. Получены уравнения, позволяющие описать зависимость соотношения ресурсных (резервных) и структурных форм биомассы от интенсивности света. Дана также зависимость содержания структурных форм в биомассе от света. Граничные световые условия находятся в диапазоне от компенсационного пункта до насыщающих значений. Ключевые слова: микроводоросли; культивирование; скорость фотосинтеза; удельная скорость роста; биохимический состав.
Введение Главным фактором, определяющим фотосинтез микроводорослей при фотоавтотрофном росте в культуре, является свет. Для количественной оценки влияния света на рост и фотосинтез микроводорослей предложено много моделей, в которых используются те или иные параметры культур. Предлагаемая работа не ставит своей целью анализ известных моделей и получаемых при этом уравнений, а в качестве основных уравнений зависимости фотосинтеза от света в данной работе используется метод линейных сплайнов. Такой выбор не случаен. В работе (Тренкеншу,1992) было показано, что на форму зависимости скорости метаболической реакции от потока фотонов влияет организация ферментных или транспортных систем в клетке, определяющих предельные скорости реакций (узкое места метаболизма). Чем лучше организована метаболическая система, тем ближе форма зависимости к идеальной, т. е. к уравнению ломаной (с насыщением), впервые предложенной Блекманом (Blackman, 1905). Еще одним доводом в пользу идеализированной модели, можно привести экспериментальные данные, полученные для мгновенных световых кривых фотосинтеза (см., например, Myers, Graham, 1971; Anning et al., 2000). Их форма намного ближе к ломаной Блекмана, чем к любым другим известным уравнениям. При описании такого рода данных линейными сплайнами значительно сужается диапазон отклонений в районе точки перегиба. Кроме того, при использовании уравнения ломаной сразу определяется граница смены лимитирующих факторов. Рассматривая влияние света на синтез биомассы микроводорослей в фотоавтотрофной культуре необходимо учитывать последовательность световых и темновых реакций, при которых ассимилируется минеральный углерод и за счет световой энергии в цикле Кальвина преобразуется в полисахариды. В свою очередь, эти первичные углеводы служат источником энергии и углеродных скелетов для всех последующих процессов биосинтеза клеточных структур. Ясно, что такая последовательность процессов преобразования минеральных веществ в структурные компоненты клеток указывает на значительные биохимические изменения в составе биомассы. Огромное количество актов биохимических преобразований предполагает их учет для построения полной динамической модели этих процессов, что приведет к огромному количеству уравнений и бессмысленности моделирования в целом. В то же время для корректного описания биохимических трансформаций при стационарном росте достаточно учесть ключевые параметры, определяющие конечные скорости процессов. Кроме того, в клетках микроводорослей можно выделить структурную часть биомассы, которая участвует в процессах биотрансформаций в качестве катализаторов. Это позволяет свести к минимуму количество уравнений.
Основные положения. Экспоненциальный рост микроводорослей в культуре возможен только в случае равенства удельных скоростей роста всех без исключения компонентов клеток. Это свойство позволяет вычленить любые формы молекулярного или макромолекулярного состава биомассы, при этом соотношения форм будут постоянными на всем промежутке экспоненциального роста. Одним из вариантов выбора таких форм может служить представление о структурных и резервных («фондовых») формах углеродных соединений у листьев высших растений и подтверждается современными данными о расходовании углеродных фондов при темновых реакциях в листьях растений (Ивлев, 2011). Эти представления можно несколько расширить, по крайней мере для микроводорослей, выделяя в клетке структурную часть биомассы и ресурсную, включающую все промежуточные соединения, из которых в дальнейшем формируется структурная часть клетки. Такой подход впервые был использован в работе (Тренкеншу, 2016 а) для количественного описания процессов трансформации макромолекулярных форм биомассы микроводорослей в темноте. Таким образом, биомасса клетки может быть представлена суммой двух основных макромолекулярных форм: структурной и ресурсной (= резервной). Перечисленные подходы являются основой предлагаемой работы.
Система уравнений. Вычленим в общей биомассе (B) клетки две основные макромолекулярные формы – структурную (Bstr) и неструктурную (=ресурсную, Bres):
Обозначим доли этих частей в биомассе:
В процессе метаболизма микроводорослей скорость синтеза биомассы будет определяться суммой скоростей синтеза структурных и накопления или расхода ресурсных составляющих:
Для культур микроводорослей невысокой плотности при полном минеральном обеспечении и без взаимного затенения клеток зависимость отношения удельной скорости синтеза продуктов световой стадии (μi) к максимальной (μm) будет определяться только величиной внешней (I0) и насыщающей (I0sat) освещенности (Тренкеншу, 2016 a):
В настоящей работе рассматриваются характеристики роста микроводорослей именно при таких световых условиях. Наиболее простой вариант динамики светозависимого роста и превращения ресурсных форм биомассы в структурные составляющие микроводорослей можно выразить в виде системы уравнений, предложенных ранее в работе (Тренкеншу, 2016 b). Скорость изменения структурных компонентов клетки при фотоавтотрофном росте определяется разностью между скоростью их синтеза из ресурсных составляющих за счет световой энергии и скоростью их естественного распада, называемого расходом на поддержание структуры (Перт, 1978). Введем обозначения: - μrs – удельная скорость трансформации ресурсных веществ в структурные, - qrs – экономический коэффициент этого перехода, - μr – удельная скорость естественного распада структур, В результате получим:
Трансформация ресурсных веществ в структурные представляет собой ферментативный процесс, который происходит в структурах клетки. Удельная скорость этого процесса может быть выражена через соотношение ресурсные/структурные формы биомассы (ε) и максимальную скорость трансформации (μmrs), достигаемую при насыщающем (εsat) или большем соотношении (Тренкеншу, 2016 a,b):
Заметим, что это уравнение является аналогом приложения концепции Друпа к лимитированию роста микроводорослей в непрерывной культуре (Droop, 1973, 1979). Окончательно, для скорости изменения структурных форм как функции соотношения форм в клетке, записываем в виде:
Скорость изменения ресурсных веществ в клетке будет определяться разностью между скоростями синтеза органического вещества в реакциях фотосинтеза и его расхода на образование структур. Для культур невысокой плотности получим:
В результате имеем систему уравнений, описывающую динамику трансформации структурных и ресурсных форм биомассы микроводорослей при фотоавтотрофном росте:
Скорость роста культуры будет представлять собой сумму скоростей изменения структурных и ресурсных форм биомассы:
Из последнего уравнения следует, что скорость роста микроводорослей зависит как от условий освещения (I0), так и от соотношения ресурсных и структурных составляющих биомассы клеток (ε). Содержание структурных и ресурсных макромолекулярных форм в биомассе микроводорослей можно выразить через их соотношение в клетках:
Непрерывная культура. Наиболее простой вариант решения полученной системы уравнений можно получить для динамически равновесного роста микроводорослей. Такой рост будет наблюдаться при непрерывном выращивании клеток, например, при поддержании плотности культуры на определенном уровне за счет слива части суспензии микроводорослей и долива такого же объема свежей питательной среды (плотностат). Плотностат отличается от хемостата тем, что скорость протока среды не задается постоянной, а определяется скоростью роста культуры, которая стабилизируется на определенном уровне, в зависимости от заданной плотности культуры. Это позволяет экспериментально измерять изучаемые характеристики с любой заданной точностью. Обозначим изучаемые величины надстрочным индексом (*), чтобы выделить их значения, полученные для такого режима выращивания:
Учитывая, что в непрерывной культуре все удельные скорости равны и равны удельной скорости роста (или протока), получим:
В настоящей работе скорость роста микроводорослей отдельно не рассматривается. А величина удельной скорости роста используется только в качестве инструмента для решения систем уравнений. Это позволяет систему уравнений упростить:
Смена лимитирующих факторов. Выделим ключевые моменты, определяющие переход от одного типа лимитирования роста к другому. Одним из них является видоспецифический параметр клеток, заложенный в виде насыщающей величины соотношения ресурсные/структурные формы органического вещества микроводорослей (εsat). Эту величину можно условно назвать ресурсным (или метаболическим) насыщением. В этой точке происходит переход от лимитирования скорости роста микроводорослей внутренней концентрацией ресурсных (резервных) веществ к ограничению скорости роста концентрацией структурных компонентов клеток, участвующих в преобразовании ресурсных веществ в структурные формы. Второй ключевой точкой является величина интенсивности света, при которой происходит переход от лимитирования скорости образования ресурсных веществ светом, к ограничению скорости концентрацией структурных компонентов клеток (световое насыщение). Эта точка также может быть выражена насыщающим параметром (ε0sat). В зависимости от соотношения насыщающих параметров искомые зависимости будут описываться разными парами уравнений. При этом насыщающие параметры служат точками сопряжения разных зависимостей. Рассмотрим случай светового лимитирования, когда при увеличении интенсивности света до насыщающих величин не происходит переключения светового лимитирующего фактора на иной. Насыщающие величины соотношений являются видоспецифическими параметрами, т.е. постоянными, это позволяет ввести коэффициент пропорциональности:
В контексте данной работы коэффициент меньше единицы.
Зависимость соотношения макромолекулярных форм биомассы от интенсивности света. Для интенсивностей света, не превышающих насыщающую, систему можно редуцировать до двух уравнений:
Определим границы применимости для полученной системы уравнений. При отсутствии света применение уравнений не имеет смысла, т.к. в темноте невозможно получение непрерывной фотоавтотрофной культуры. Такой случай подробно рассмотрен в работе (Тренкеншу, 2016 b). Рост микроводорослей в фотоавтотрофной культуре возможен только при интенсивности света выше определенной величины, называемой компенсационным пунктом фотосинтеза. Световой компенсационный пункт. Этой точке на световой кривой роста будут соответствовать минимальные значения соотношения ресурсные/структурные формы биомассы. Скорость роста будет равной нулю:
Из уравнений находим величину соотношения ресурсные/структурные формы биомассы в точке, соответствующей световому компенсационному пункту при фотоавтотрофном росте микроводорослей. В этом случае величина соотношения будет минимальной для непрерывной культуры. Введем для удобства безразмерную величину интенсивности света:
Из первого уравнения системы находим соотношение ресурсные/структурные формы биомассы в световой компенсационной точке: Содержание структурных форм органического вещества в биомассе клеток в световой компенсационной точке будет максимальным. Вводя соответствующие обозначения, получим:
Решение системы позволяет не только найти зависимость величины светового компенсационного пункта от минимальной величины соотношения ресурсных и структурные форм, но и выразить ее через кинетические коэффициенты:
Полученная величина интенсивности света является нижней границей возможности использования системы уравнений для описания светозависимого роста. Верхней границей возможности использования системы уравнений является насыщающая интенсивность света. При насыщающей интенсивности света система уравнений принимает вид:
Решение последней системы уравнений показывает, что максимальное значение искомого соотношения определяется только кинетическими константами:
Положительные значения будут наблюдаться только при выполнении следующего условия соотношения между кинетическими коэффициентами:
В противном случае рост микроводорослей не будет лимитирован светом.
При максимальном соотношении ресурсных и структурных форм доля структурных макромолекулярных составляющих биомассы будет минимальной:
Окончательно система уравнений с граничными световыми условиями запишется в виде:
Решая систему уравнений, находим зависимость соотношения ресурсных и структурных макромолекулярных форм в биомассе микроводорослей при экспоненциальном росте от интенсивности света:
Содержание структурных форм в биомассе можно получить из уравнения:
Графические представления. Учитывая, что кинетические коэффициенты являются видоспецифическими, что существенно влияет на форму искомой зависимости, выразим их и соотношения макромолекулярных форм биомассы в безразмерном виде. Экономические коэффициенты метаболических реакций уже заданы в безразмерном виде. Примем, что эффективность преобразования ресурсных веществ в структурные формы составляет 40% (qrs = 0,4). Выразим удельные скорости процессов в безразмерных величинах, отнесенных к максимальной удельной скорости преобразования ресурсных составляющих биомассы в структурные. Считаем, что максимальная удельная скорость преобразования ниже скорости фотосинтеза на 20%, а скорости поддержания – в 20 раз выше:
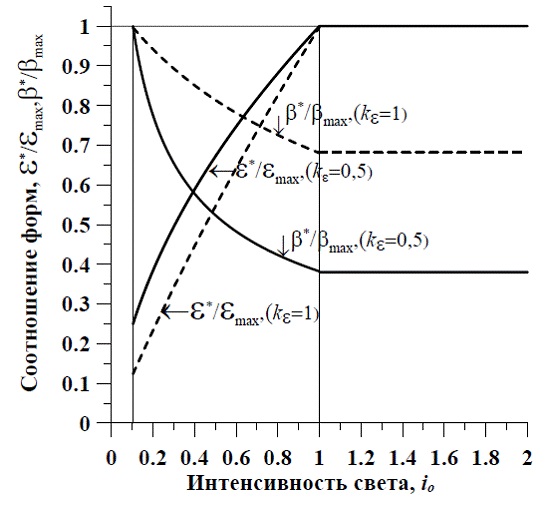
Максимальное значение соотношения ресурсные/структурные формы биомассы можно принять за нормирующий множитель и выражать полученные зависимости в долях от максимального значения. Существенное влияние на состав биомассы оказывает соотношение насыщающих величин, т.к. эти параметры являются видоспецифическими. На рис. 1 для примера показаны зависимости соотношения ресурсные/структурные формы макромолекулярного состава биомассы и содержания структурных форм в биомассе микроводорослей от интенсивности света. Расчет сделан для непрерывных культур невысокой плотности, с использованием коэффициентов, приведенных выше для двух случаев: kε=1 и kε=0,5.
Рис. 1. Зависимость соотношения ресурсных и структурных форм (ε*=Bres/Bstr) и содержания структурных компонентов в биомассе микроводорослей (β*=Bstr /B) от интенсивности света в непрерывной культуре низкой плотности Fig. 1. The dependence of the ratio of resource and structural forms (ε*=Bres/Bstr) and content of structural components in biomass of microalgae (β*=Bstr /B) on light intensity in continuous culture of low density
Вывод Показано, что в динамически равновесной (непрерывной) культуре микроводорослей макромолекулярный состав биомассы клеток зависит от поверхностной освещенности культуры. Получены уравнения и граничные условия, позволяющие описать зависимость соотношения ресурсных (резервных) и структурных форм биомассы и содержания структурных компонентов в биомассе от интенсивности света в диапазоне от компенсационного пункта до насыщающих значений.
Работа выполнена в рамках гос. задания № 1001-2014-0017 «Разработка научных основ решения гидробиологических и биотехнологических проблем интегрированного управления прибрежными зонами».
Список литературы 1. Ивлев А.А. Колебательная природа углеродного метаболизма в фотосинтезирующей клетке по данным изотопного состава углерода // Успехи современной биологии. 2011. № 2 (131). С. 178–192. 2. Перт С.Дж. Основы культивирования микроорганизмов и клеток. М., 1978. – 333 с. 3. Тренкеншу Р.П. Применение теории массового обслуживания в биокинетике // Эволюционное моделирование и кинетика. – Новосибирск: Наука, 1992. – С. 125–160. 4. Тренкеншу Р.П. Модификация уравнений динамики макромолекулярного состава микроводорослей // Морские биологические исследования: достижения и перспективы. – Севастополь: Экоси-гидрофизика, 2016 a. – С. 448–451. 5. Тренкеншу Р.П. Динамическая модель биотрансформации резервных и структурных форм биомассы микроводорослей в темноте // Вопросы современной альгологии. 2016 b. №2(12). URL: http://algology.ru/967 6. Anning T., MacIntyre H. L., Pratt S.M., Sammes P.J., Gibb S., Geider R.J. Photoacclimation in the marine diatom Skeletonema сostatum // Limnology & Oceanography. 2000. 45. P. 1807–1817. 7. Blackman F.F. Optima and limiting factors // Ann. Bot. Lond. 1905. 19. P. 281–295. 8. Droop M.R. Some thoughts on nutrient limitation in algae // J. Phycolog. 1973. 9. P. 264–272. 9. Droop M.R. The definition of X and Q in the cell quota model // J. Exp. Mar. Biol. Ecol. 1979. 39. P. 203. 10. Myers J., Graham J. The Photosynthetic Unit in Chlorella Measured by Repetitive Short Flashes // Plant Physiol. 1971. 48. P. 282–286. Статья поступила в редакцию 31.10.2017
Influence of light on macromolecular composition of microalgae Rudolf P. Trenkenshu It is shown that in dynamically equilibrium (continuous) culture of low density macromolecular composition of microalgae biomass depends on the surface light. Equations allowing to describe the ratio dependence of resource (backup) and structural forms of biomass on light intensity. The dependence of the content of structural forms in the biomass on the light is also given. Boundary light conditions are within the range of compensation point and saturating values. Key words: microalgae; cultivation; photosynthesis rate; specific growth rate; biochemical composition.
Об авторе Тренкеншу Рудольф Павлович – Trenkenshu Rudolf P. кандидат биологических наук r.trenkenshu@rambler.ru Корреспондентский адрес: Россия, 299011, Севастополь, пр. Нахимова, 2, ФГБУН ИМБИ; тел. (869)-2550795.
ССЫЛКА НА СТАТЬЮ: Тренкеншу Р.П. Влияние света на макромолекулярный состав микроводорослей в непрерывной культуре невысокой плотности (Часть 1) // Вопросы современной альгологии. 2017. № 2 (14). URL: http://algology.ru/1180 Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно.
При перепечатке ссылка на сайт обязательна
На ГЛАВНУЮ
|
|||
|
| ||