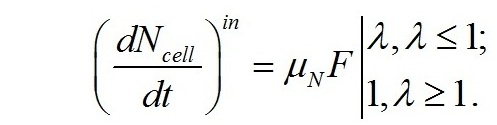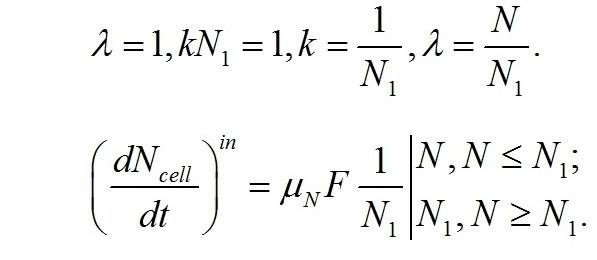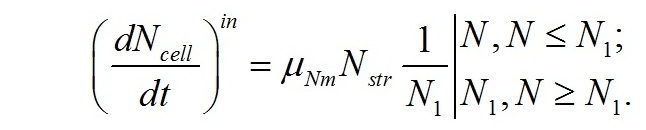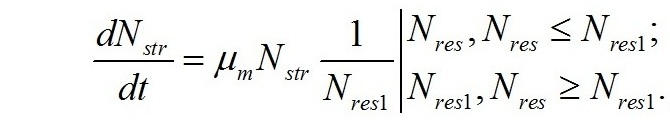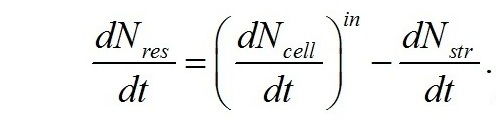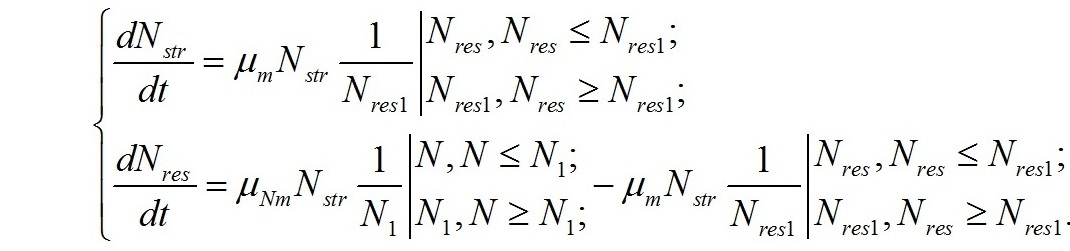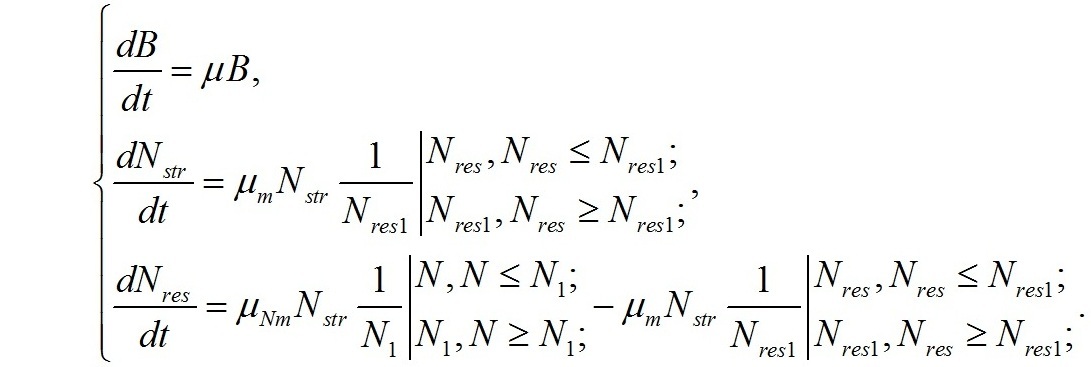|
|
|
|
по Материалам Международной конференции «Экологическая физиология водных фототрофов: распространение, запасы, химический состав и использование» VI Сабининские чтения 1 ноября - 15 декабря 2017 г. Модель трансформации форм азотистых соединений клетками микроводорослей
Тренкеншу Р.П., Лелеков А.С. Rudolf P. Trenkenshu, Alexander S. Lelekov
Институт морских биологических исследований им. А.О.Ковалевского (Севастополь, Россия)
УДК 579.017.8:57.036
Предложена динамическая модель трансформации форм азотистых соединений в клетках микроводорослей. Внутриклеточная концентрация азота рассматривается в виде суммы структурных и резервных соединений. Скорость поступления азота в клетку и скорость его расхода на синтез структурной составляющей заданы в виде линейных сплайнов. Получена система дифференциальных уравнений позволяющая описать не только соотношения различных форм азота в клетке, но и их зависимость от его концентрации в среде. Ключевые слова: микроводоросли; субстрат-зависмый рост; азот; математическая модель.
Введение В последние годы резко возросло количество публикаций по изучению влияния азота на рост и биохимический состав микроводорослей в природе и культуре. Это связано с множеством национальных программ по поиску возобновляемых источников энергии, одним из которых рассматриваются микроводоросли. Причем повышенное внимание уделяется их углеводному и липидному составу, основному сырью для производства биотоплива. Выяснилось, что доля углеводов и жиров в клетках микроводорослей возрастает при дефиците азота в среде (Боровков, Гудвилович, 2017; Fidalgo, 1995; Hu, 2008; Sánchez-Saavedra, 2017; Silva, Sforza, 2017). В это же время стала бурно развиваться аквакультура ценных пород рыб, появилась потребность в пигментах и полиненасыщенных жирных кислотах (имеющих микроводорослевое происхождение), которые стимулируют рост и выживаемость личинок рыб, повышают качество конечной продукции (Gatenby et al., 2003). Вместе с тем, медицинские исследования показали важную роль полиненасыщенных жирных кислот для здоровья человека, особенно, w3 и w6 кислот (Cartens, 1996; Desbois, 2008). Наряду с успешным решением ряда задач по техническому обеспечению процессов массового культивирования микроводорослей, наличием современного инструментария для оценки макромолекулярного состава получаемой биомассы и др., основной проблемой остается отсутствие полноценной количественной теории управления фотобиосинтезом микроводорослей. В настоящее время имеются все предпосылки для создания такой теории. Накоплены знания о молекулярных механизмах световых и темновых процессов фотобиосинтеза. Найдены основные метаболические пути синтеза органического вещества в клетках микроводорослей, изучены системы транспорта веществ в клетку и их формирование. Разработаны принципы и подходы к возможности использования формальных приемов для создания математических моделей метаболических процессов биосинтеза и их связи с внешними условиями среды (Тренкеншу, Лелеков, 2017; Тренкеншу, Лелеков, Новикова, 2017). Учитывая большое количество стадий трансформации минеральных веществ в органические молекулы, при моделировании процессов преобразования математическое описание будет содержать огромное количество дифференциальных уравнений, что приводит к бессмысленности моделирования в целом. Однако пользуясь принципом минимума, можно редуцировать большую систему до нескольких уравнений. Для этого нужно знать узкое место в метаболической цепи ферментативных и транспортных реакций. Причем это узкое место в большинстве случаев не обязательно знать точно, в кинетических моделях его можно выразить через максимальные скорости процессов, которые можно измерить непосредственно (Тренкеншу, 2016; Тренкеншу, 2017; Тренкеншу, Лелеков, Новикова, 2017). Следующим, после углерода, минеральным биогеном, который может влиять на скорость роста и макромолекулярный состав биомассы микроводорослей, является азот. В предлагаемой работе рассматривается влияние концентрации азота в среде на кинетику роста и трансформацию структурных и резервных форм азота в культуре и клетках микроводорослей. Кинетика поглощения азота клеткой. Рассмотрим скорость поступления растворенного в питательной среде азота (концентрация N в среде) в клетки микроводорослей. Азот, находящийся в клетках, рассматриваем в виде двух основных форм: структурной и резервной. К структурной части относим азот, входящий в структуру клеточной массы (Nstr). Эта часть биомассы в основном представлена белками. Сюда же относим и небелковые, но входящие в структуры формы азота (например, коферменты). Резервная часть азота в клетках (Nres) представлена той частью, которая еще не вошла в структурные формы и рассматривается как субстрат в последовательности метаболических реакций (включая интермедиаты) (Perez-Garcia, 2010). Такое представление полностью согласуется с концепцией Друпа о том, что скорость роста микроорганизмов зависит от внутриклеточной концентрации лимитирующего субстрата (Droop, 1973). Внутриклеточная концентрация азота Ncell (отнесенная к объему культуры), равна сумме:
Общий баланс азота в культуре:
где N0 – начальная концентрация азота в среде. Последнее выражение, несмотря на очевидную простоту, отражает фундаментальный закон сохранения азота в закрытой системе: на рис. 1 представлена динамика концентрации нитратного азота в среде и структурного азота при накопительном культивировании Dunaliella salina.
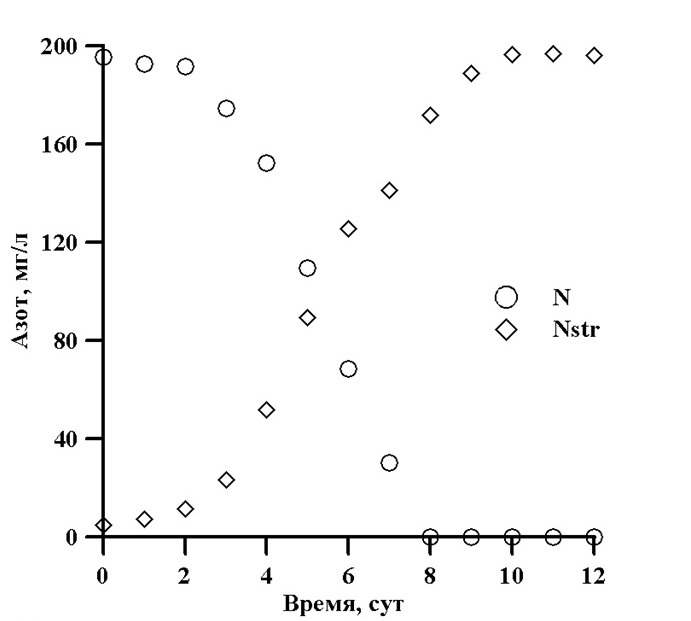
Рис. 1. Динамика концентрации нитратного азота в среде и структурного азота при культивировании Dunaliella salina (Лелеков, 2007; Лелеков, Гудвилович, 2010)
Структурный азот рассчитан делением концентрации белка на 6,25 (Ho, 2012). Из представленных на рисунке данных видно, что на восьмые сутки роста культуры концентрация азота в среде стала близкой к нулю, но рост продолжался еще двое суток, вплоть до исчерпания внутриклеточного резерва. Весь азот в нитратной форме, с точностью до 0,5%, к моменту достижения максимальной биомассы превратился в структурный. Скорость поступления азота в клетки будет определяться количеством транспортных систем (F), их активностью (μN) и приведенной плотностью потока молекул (λ) из внешней среды (Тренкеншу, 2010):
Приведенная плотность потока молекул субстрата, попадающих в клетки за счет диффузии или транспортной системы клеток, пропорциональна (коэффициент k) концентрации данного вещества во внеклеточной среде культуры:
При некоторой концентрации N1 в среде, поток становится единичным, и величину приведенной плотности потока азота в клетку можно выразить через эту величину, а уравнение для скорости поступления азота в клетку записать в виде:
Учитывая, что транспортные системы относятся к структурным компонентам клетки, их количество можно выразить через концентрацию структурных форм азота (fN=F/Nstr). Величина
Кинетика трансформации форм азота внутри клетки. Скорость синтеза структурной составляющей биомассы и, соответственно, расхода резервного азота будет зависеть от его концентрации, активности (msin) и количества ключевого фермента (выраженного через долю структурного азота fsin), контролирующего скорость образования структурных форм биомассы. Произведение msinfsin=mm представляет собой максимальную удельную скорость включения азота в структуру клетки или синтеза структурных компонентов. В итоге имеем:
Скорость изменения концентрации резервного азота будет равна разности между скоростью поступления азота в клетку и скоростью его расхода на синтез структур:
В результате имеем систему уравнений, описывающую динамику внутриклеточных переходов одних форм азота в другие:
Используя полученную систему уравнений можно оценить не только ростовые характеристики микроводорослей, но и решить обратную задачу – найти зависимость форм азотистых соединений в биомассе клеток от внешних концентраций азота в среде и скорости их роста. Дополним систему уравнений ростовой характеристикой микроводорослей. Если μ – удельная скорость роста, а В – плотность культуры выраженная концентрацией клеток или концентрацией биомассы, то скорость роста:
В результате имеем:
Фактически, полученная система уравнений является искомой динамической моделью роста и трансформации структурных и резервных форм азотистых соединений в клетках микроводорослей в дифференциальной форме. Решение системы при заданных условиях позволит получить интегральные зависимости концентрации различных форм азота от времени, а значит, даст возможность рассчитать кинетические характеристики μNm, μm, которые являются видоспецифическими коэффициентами.
Заключение В данной работе предложена математическая модель, которая является логическим продолжением наших предыдущих исследований кинетики ассимиляции азота клетками микроводорослей. Введенная ранее величина истинной потребности микроводорослей в лимитирующем субстрате (Тренкеншу, Лелеков, 2017) есть динамическая характеристика, которая изменяется в широком диапазоне в зависимости от потока азота из внешней среды. Эти изменения можно объяснить, разделив внутриклеточный азот на структурный и резервный: в условиях недостатка азота его внутриклеточный пул (резервная составляющая) будет исчерпан. Предлагаемая математическая модель позволяет также описать процесс синтеза белка в условиях полного отсутствия азота во внешней среде.
Работа выполнена в рамках госзадания по НИР «Разработка научных основ решения гидробиологических и биотехнологических проблем интегрированного управления прибрежными зонами» № АААА-А18-118021350003-6
Список литературы 2. Лелеков А.С., Горбунова С.Ю., Боровков А.Б. Динамика азота и фосфора в среде при интенсивном культивировании микроводоросли Dunaliella salina // Экология моря. 2007. Вып. 74. С. 21–24. 3. Лелеков А.С., Гудвилович И.Н. Продукционные характеристики роста и биосинтеза квазинепрерывной культуры зелёной микроводоросли Dunaliella salina Teod. // Экология моря. 2010. Спец. вып. 80: Биотехнология водорослей. C. 59–66. 6. Тренкеншу Р.П., Лелеков А.С. Моделирование роста микроводорослей в культуре. – Севастополь: ООО «Константа-Принт», 2017. – 152 с. 7. Тренкеншу Р.П., Лелеков А.С., Новикова Т.М. Линейный рост морских микроводорослей в культуре // Морской биологический журнал. 2017. Т.2, №4. (В печати). 9. Desbois A.P., Lebl T., Yan L., Smith J.V. Isolation and structural characterisation of two antibacterial free fatty acids from the marine diatom Phaeodactylum tricornutum // Appl. Microbiol. Biotechnol. 2008. V.81. Р. 755–764. 10. Droop M.R. Some thoughts on nutrient limitation in algae // J. Phycol. 1973. V.9. P. 264–272. 11. Fidalgo P., Cid A., Abalde J., Herrero C. Culture of the marine diatom Phaeodactylum tricornutum with different nitrogen sources: Growth, nutrient conversion and biochemical composition // Cah. BioI. Mar. 1995. V.36. P. 165–173. 13. Ho S., Chen C., Chang J. Effect of light intensity and nitrogen starvation on CO2 fixation and lipid/carbohydrate production of an indigenous microalga Scenedesmus obliquus CNW-N // Bioresource Technology. 2012. V.113. P. 244–252. 14. Hu Q., Sommerfeld M., Jarvis E., Ghirardi M. et al. Microalgal triacylglycerols as feedstocks forbiofuel production: perspectives and advances // J. Plant. 2008. V.54, №4. P. 621–639. 16. Sánchez-Saavedra M. P., Castro-Ochoa F. Y., Nava-Ruiz V. M. et al. Effects of nitrogen source and irradiance on Porphyridium cruentum // J. Appl. Phycol. 2017. https://doi.org/10.1007/s10811-017-1284-2. 17. Silva F., Sforza E. Stability of carbohydrate production in continuous microalgal cultivation under nitrogen limitation: effect of irradiation regime and intensity on Tetradesmus obliquus // J. Appl. Phycol. 2017. https://doi.org/10.1007/s10811-017-1252-x. Статья поступила в редакцию 28.11.2017 .
Model of transformation of the nitrogen compounds by microalgae cells Rudolf P. Trenkenshu, Alexander S. Lelekov Kovalevsky Institute of Marine Biological Research of RAS (Sevastopol, Russia) Dynamic model of transformation of forms of nitrogen compounds in microalgae cells has been proposed. Intracellular concentration of nitrogen is considered as the sum of the structural and reserve compounds. The rate of nitrogen flow into the cell and the rate of its use for the synthesis of structural component has been described as linear splines. The system of differential equations allowing to describe not only ratios of different forms of nitrogen in the cell but also their dependence on nitrogen concentration in the medium has been obtained. Key words: microalgae; substrate dependent growth; nitrogen; mathematical model.
Об авторах Тренкеншу Рудольф Павлович – Trenkenshu Rudolf P. кандидат биологических наук r.trenkenshu@rambler.ru Лелеков Александр Сергеевич – Lelekov Alexander S. кандидат биологических наук a.lelekov@yandex.ru Корреспондентский адрес: Россия, 299011, Севастополь, пр. Нахимова, 2, ФГБУН ИМБИ; тел. (869)-2550795.
ССЫЛКА НА СТАТЬЮ: Тренкеншу Р.П., Лелеков А.С. Модель трансформации форм азотистых соединений клетками // Вопросы современной альгологии. 2018. № 1 (16). URL: http://algology.ru/1247 Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно. При перепечатке ссылка на сайт обязательна
К разделу ОБЗОРЫ, СТАТЬИ И КРАТКИЕ СООБЩЕНИЯ

|
|||
|
| ||