|
|
|
|
29 ноября - 29 декабря 2014 г. Морфометрические показатели и химический состав Saccharina japonica, культивируемой в Японском море (Приморский край)
Аминина Н.М., Гурулева О.Н. Natalia M. Aminina, Olga N. Guruleva
Тихоокеанский научно-исследовательский рыбохозяйственный центр (ТИНРО-Центр)
УДК 577.1:582.272 (571.63)
Показана сезонная изменчивость морфометрических показателей слоевищ культивируемой сахарины японской, а также химического состава как целого слоевища, так и отдельных его частей. Накопление максимальной длины и массы слоевища у двухлетних водорослей приходится на июнь. В этот же период отмечено увеличение концентрации белка и минеральных веществ в талломах. Наибольшее содержание альгиновой кислоты обнаружено в июле-августе. С мая по июнь больше всего органических веществ определяется в верхней части пластины, в августе же их, особенно белка, накапливается больше в нижней части растения. Маннита, и альгиновой кислоты больше определяется в зоне интенсивного биосинтеза – по краям слоевища; фукоидана – в центральной части. Установлено, что содержание фукоидана и альгиновой кислоты увеличивается в зоне образования спороносной ткани. Ключевые слова: бурые водоросли; Saccharina japonica; морфометрические показатели; период вегетации; спороносная ткань; химический состав; альгиновая кислота; манит; фукоидан; минеральные вещества; белок.
У водорослей, как и у наземных растений, имеются определенные фенологические закономерности роста и развития. Свет и температура считаются определяющими факторами, влияя на интенсивность фотосинтеза, развитие спор, нарастание массы талломов. В дальневосточных морях интенсивный рост растений наблюдается в весенние месяцы. Летом темп роста снижается, а к концу лета чаще всего начинается разрушение слоевищ растений. В большинстве случаев рост водорослей коррелирует одновременно с освещенностью и температурой (Ярцева и др., 1977). Другие исследователи считают, что рост слоевищ ламинариевых в большей степени зависит от температуры воды (Шпакова, 1985). Причем наиболее значимыми являются колебания температуры в весенние месяцы (Кулепанов В.Н., Дзизюров В.Д., Жильцова Л.В., 2002). Сезонное изменение температуры воды формирует течение физиологических процессов морских растений, определяет репродуктивные функции. Сезонная зависимость характерна и для процессов метаболизма водорослей. Свет различной интенсивности определяет форму хлоропластов и степень фотосинтеза (Титлянов и др., 1975, 1978). Соответственно, меняется интенсивность метаболизма, состав и количество синтезируемых веществ. Известно, что в ламинариевых водорослях, обитающих в морях Дальнего Востока, весной накапливается минеральных веществ больше, чем в летние месяцы. Количество органических веществ, наоборот, увеличивается в летние месяцы. При прекращении роста и начале деградационных процессов их соотношение снова меняется, уменьшается обводненность тканей талломов (Камнев, 1999). От возраста растения и стадии его развития зависит состав органических веществ. Различия химического состава характерны и для отдельных частей слоевища, выполняющих разные физиологические функции, например, пластины, черешков и ризоидов. Известно, например, что в период развития спороносной ткани изменяется соотношение аминокислот, при этом увеличивается количество глутаминовой кислоты. Отличия наблюдаются и по топографии пластины водорослей. Так, удельная поверхность пластины и уровень фотосинтетической деятельности выше в краевой зоне ламинариевых водорослей. Соответственно, в этой части должны активнее протекать метаболические реакции и отличаться химический состав. Установлено, например, что в краевой и центральной зонах пластины ламинариевых различается соотношение минеральных и органических веществ (Камнев, 1999). У представителей семейства ламинариевых развитие поверхности пластины имеет сезонно выраженный квазипериодический характер. В течение года, особенно в весенне-летний период, меняется физиологическая активность пластины и ее отдельных частей, соответственно, должен отличаться и химический состав этих частей. Целью нашего исследования было определение взаимосвязи между основными морфометрическими показателями и химическим составом ламинарии японской в период с мая по август, как в целой пластине, так и в отдельных ее частях. Материалы и методы В качестве объекта исследований использовали слоевища культивируемой бурой водоросли сахарины японской (Saccharina japonica = Laminaria japonica), которые были заготовлены с мая по август 2012 г. на плантации в б. Анна (южное Приморье). Для определения компонентов по топографии пластины ее разрезали на 3 равные части (верх, середина, низ), а также разделяя центральную и краевые части пластин. Для проведения химических анализов слоевища водорослей и их отдельные части высушивали и измельчали до кусочков размером 0,1-0,5 мм, подготавливали среднюю пробу, которую хранили в банке с притертой пробкой. Образцы водорослей исследовали на содержание воды, золы, альгиновой кислоты стандартными методами (ГОСТ 26185-84). Общее содержание азотистых веществ определяли микрометодом по Кьельдалю на анализаторе азота «Kjeltec auto» 10 SO Analyzer (Tecator, Япония). Количество фукозы – спектрофотометрически по цветной реакции с L-цистеином и серной кислотой (Dische et al., 1948; Усов и др., 2001). Результаты и обсуждение Считается, что в Приморье наиболее благоприятные условия для роста и развития ламинариевых водорослей начинаются с конца марта – начала апреля и продолжаются до середины июля при температурах 1-15ºС (Гусарова, 1988). Максимальный темп роста наблюдается в апреле-мае (3-6 см в сутки), летом водоросли достигают своих максимальных размеров. Популяция ламинарии японской на юге Приморья (от м. Поворотного до зал. Ольги) характеризуется активным накоплением массы, ранней остановкой вегетативного роста, коротким периодом развития спороносной ткани по сравнению с популяцией более северных районов (Иванова, Гусарова, 2005). Условия развития сахарины японской на плантации в южной части Приморья (бухта Анна) отличаются от условий произрастания естественных зарослей по глубине и способу прикрепления слоевищ. Это, естественно, может менять течение физиологических процессов и влиять на темпы роста водорослей. Исследования показали, что основные закономерности развития сахарины японской при ее плантационном выращивании сохраняются. Интенсивное увеличение размеров и массы слоевищ культивируемых водорослей наблюдается весной (рис. 1).
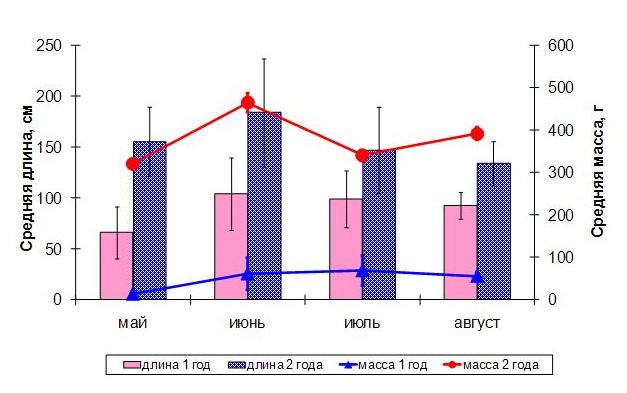
Рис. 1. Средняя длина и масса пластин сахарины японской в разные периоды вегетации.
Максимальная длина водорослей приходится на июнь первого и второго года развития (100 и 180 см соответственно). По накоплению массы растения первого и второго года несколько отличаются. Максимальная масса у двухлетних водорослей приходится на июнь, в июле она уменьшается, а в августе снова увеличивается. Масса однолетних водорослей мало отличается в летние месяцы, но в среднем она выше в июле. Изменения морфометрических показателей водорослей связаны с их метаболизмом, что, соответственно, отражается на их химическом составе. Для ламинариевых водорослей характерна высокая обводненность тканей пластин в периоды их наиболее интенсивного роста, особенно весной (Королева, 2004). Ранее было установлено, что по мере роста пластины ламинариевых водорослей возрастает обводненность тканей, в сухом остатке постепенно возрастает количество органических веществ (Камнев, 1999). При замедлении и прекращении ростовых процессов содержание органических веществ в пластине уменьшается, а минеральных – увеличивается. Наши исследования подтверждают, что в однолетних водорослях содержание минеральных веществ выше, а органических – ниже, чем в двухлетних водорослях (табл. 1). Это соотношение меняется в течение года: в весенний период интенсивный рост растений связан с преобладанием минеральных веществ, замедление темпов роста летом сопровождается увеличением суммы органических веществ.
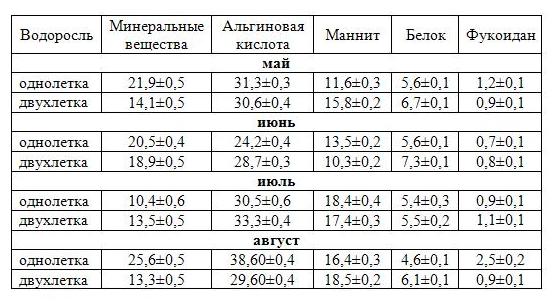
Таблица 1. Химический состав сахарины японской в разные периоды вегетации (% на сухое вещество)
В однолетней культивируемой сахарине японской максимальное количество минеральных веществ, а также белка, накапливается в мае. Второй максимум накопления минеральных веществ в слоевище наблюдается в августе, в это же время отмечено наибольшее количество полисахаридов. Для двухлетних водорослей характерна несколько иная картина: наибольшее количество минеральных веществ и белка определено в июне, альгиновой кислоты – в июле. Вероятно, у водорослей разного возраста смещены по времени физиологические процессы, в частности спорообразование, которое у двухлетних водорослей протекает позже, после чего наблюдается разрушение пластин. Считается, что с возрастом содержание альгинатов в бурых водорослях увеличивается (Камнев, 1999). Однако при анализе культивируемой ламинарии больше всего полисахаридов определено у однолетних водорослей, а не у двухлетних. Возможно, это связано с различной степенью экстракции альгинатов, которая, как известно, зависит от структурного состава полисахаридов в растениях разного возраста. При сравнении тенденций изменения химического состава и морфометрических характеристик сахарины японской установлено, что максимальная длина и масса у двухлетних растений в июне совпадает с максимальным содержанием белка и минеральных веществ (рис. 2).
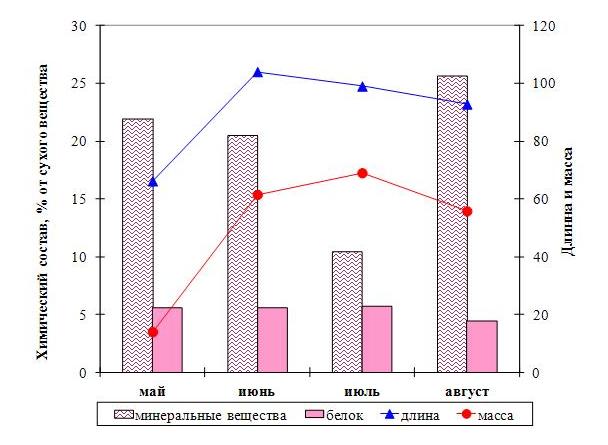
Рис. 2. Изменение химического состава и морфометрических показателей у двухлетней сахарины японской в разные периоды вегетации.
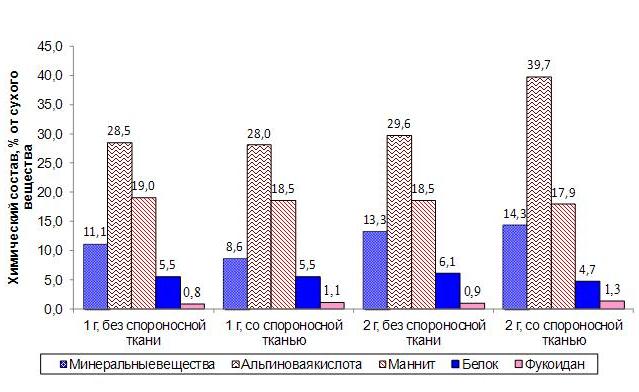
У однолетних водорослей процессы протекают иначе. Максимальная длина и масса в июне-июле совпадает с максимальным количеством белка и маннита. Количество минеральных веществ в это время уже уменьшается, а альгиновой кислоты только начинает увеличиваться. Таким образом, взаимосвязь между морфометрическими показателями и химическим составом имеет ярко выраженный сезонный характер. В мае-июне значительное увеличение длины и массы водорослей сопровождается накоплением минеральных веществ и белка. При изменении синтеза в сторону накопления полисахаридов происходит уменьшение морфометрических показателей. Ранее было установлено, что в спорофиллах водорослей, относящихся к роду алария и ундария, может накапливаться значительное количество фукоидана по сравнению с пластиной (Усов и др., 2005, Skriptsova et al., 2012). Предполагается, что повышенное содержание фукоидана необходимо для нормального функционирования спорообразующих тканей. В связи с этим мы попытались найти отличия в составе участков пластины сахарины японской, различающихся по наличию спороносной ткани. Образование спороносной ткани у однолетней сахарины японской начинается в июле, у двухлетней в августе. У двухлетних водорослей в зоне образования спороносной ткани повышается содержание фукоидана и альгиновой кислоты, тогда как количество маннита и белка – снижается (рис. 3). У однолетних водорослей концентрация этих соединений в слоевищах со спороносной тканью и без нее мало различается.
Рис. 3. Изменение химического состава сахарины японской в процессе спороношения.
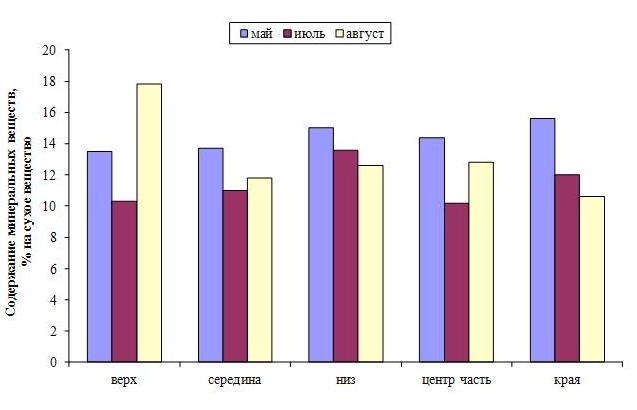
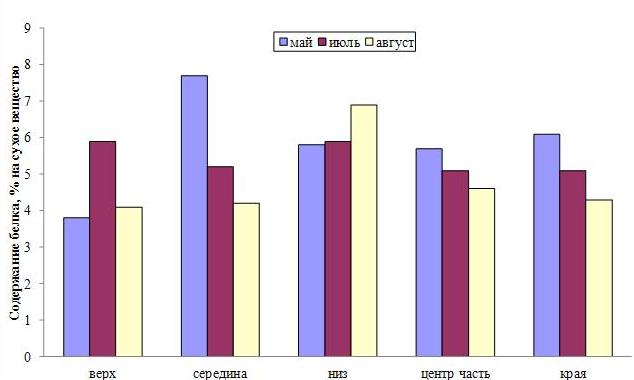
Ранее было показано, что функциональная активность ламинариевых водорослей, а следовательно и их химический состав, отличаются в разных частях пластины (Камнев, 1999). Например, в центральной зоне пластины общее содержание органических веществ более высокое по сравнению с краевой зоной, для минеральных веществ наблюдается обратная картина. Наши исследования показали, что это соотношение, а также соотношение отдельных органических веществ меняется в течение сезона. В мае-июне минеральных веществ содержится больше всего в нижней половине пластины, а также по краям (рис. 4). В августе происходит перераспределение минеральных веществ с увеличением их содержания в верхней части пластины, причем в ее центре. Белок же концентрируется в центре, но в нижней части пластины (рис. 5).
Рис. 4. Распределение минеральных веществ по топографии слоевища двухлетней сахарины японской в разные периоды вегетации.
Рис. 5. Распределение белка по топографии слоевища двухлетней сахарины японской в разные периоды вегетации.
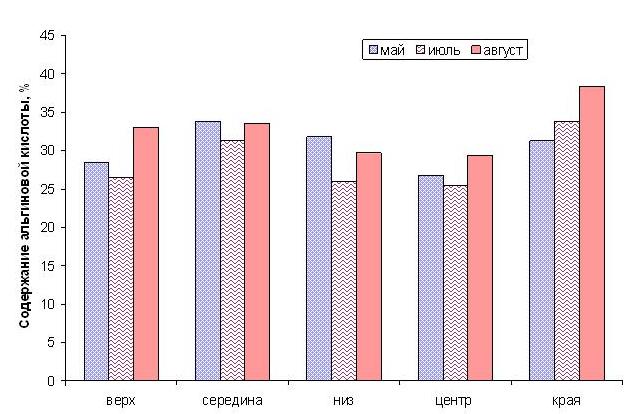
Альгиновой кислоты больше содержится в средней и в краевых частях пластины во все исследованные месяцы (рис. 6). Самое высокое содержание альгиновой кислоты в конце лета отмечено в месте расположения спороносной ткани.
Рис. 6. Распределение альгиновой кислоты по топографии слоевища двухлетней сахарины японской в разные периоды вегетации.
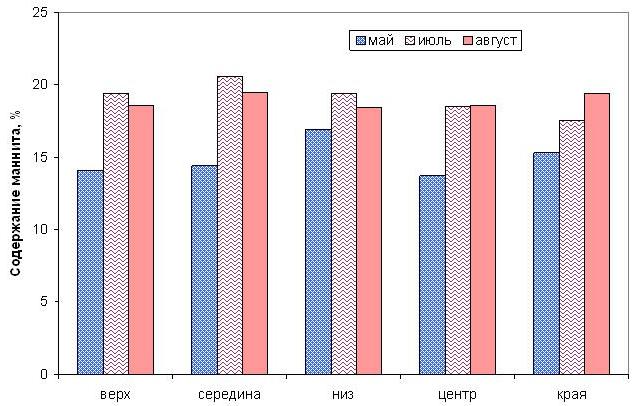
Распределение маннита в пластине в мае совпадает с накоплением минеральных веществ: больше всего его содержание в нижней половине, причем по краям пластины (рис. 7). В июле и августе маннит концентрируется в центральной части слоевища.
Рис. 7. Распределение маннита по топографии слоевища двухлетней сахарины японской в разные периоды вегетации.
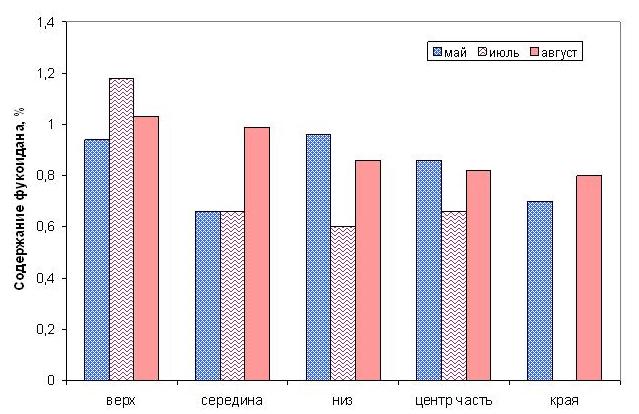
Распределение фукоидана по частям слоевища также варьирует. В мае, например, фукоидан концентрируется в нижней и в центральной части слоевища, в июле – в верхней части и по краям пластины, августе – в верхней и средней части (рис. 8).
Рис. 8. Распределение фукоидана по топографии слоевища двухлетней сахарины японской в разные периоды вегетации.
Анализ химического состава был проведен не только в пластине, но и в органах прикрепления сахарины японской. Минеральных веществ, альгиновой кислоты и белка больше всего в ризоидах, маннита и фукоидана – в пластине. Черешок по содержанию минеральных веществ и белка мало отличается от пластины, а по количеству маннита – от ризоидов. Интересен тот факт, что в черешке обнаружено фукоидана больше, чем в пластине. Заключение В ходе роста и развития Saccharina japonica изменения морфометрических показателей и химического состава носят выраженный сезонный характер. Это характерно как для целого слоевища, так и для отдельных его частей. Наблюдаются различия в темпах роста и накоплении минеральных и органических веществ между растениями разного возраста. В целом, интенсивный рост водорослей весной сопровождается накоплением минеральных веществ и усиленным синтезом белка. Летом темпы роста падают и метаболизм меняется в сторону накопления углеводов. В мае-июле минеральных веществ больше всего концентрируется в нижней части слоевища, причем по краям таллома. В августе, в период спороношения двухлетних водорослей, распределение веществ в пластине водоросли меняется. Краевая часть содержит теперь больше органических веществ, а минеральные вещества концентрируются в верхней части слоевища. У двухлетних водорослей в зоне образования спороносной ткани значительно повышается содержание альгиновой кислоты и фукоидана, а маннита и белка – снижается. У однолетних водорослей концентрация этих соединений в слоевищах со спороносной тканью и без нее мало различается. Следовательно, в процессе роста и развития Saccharina japonica наблюдаются значительные изменения метаболизма пластины и ее отдельных частей. Это свидетельствует о значительной функциональной изменчивости ламинариевых водорослей и их адаптационной способности к изменяющимся условиям окружающей среды.
Список литературы 1. ГОСТ 26185–84. Водоросли морские, травы морские и продукты их переработки. Методы анализа. – М.: Издательство стандартов, 1984. – 53 с. 2. Гусарова И.С. Сублиторальная растительность и ее сезонная динамика в одной из бухт северо–западной части Японского моря // Гидробиологические исследования заливов и бухт Приморья. Владивосток: ДВНЦ АН СССР, 1984. C. 12–27. 3. Иванова Н.В., Гусарова И.С. Морфологическая дифференциация Laminaria japonica Aresch. в сублиторали северного приморья // Комаровские чтения. Владивосток:Дальнаука, 2005. Вып. 51. С. 198-209. 4. Камнев А.Н. Возрастная морфофизиология бурых водорослей (на примере представителей порядков фукусовых и ламинариевых). Автореф. дис. на соиск. уч. ст. док. биол. наук. – М., 1999. – 74 с. 5. Королева Т.Н. Развитие бурой водоросли Laminaria bongadiana Р. еt R. в прикамчатских водах: Автореф. дис.. канд. биол. наук. – М., 2004. – 28 с. 6. Кулепанов В.Н., Дзизюров В.Д., Жильцова Л.В. Факторы, определяющие динамику запасов ламинарии японской у побережья Приморья // Приморье – край рыбацкий: материалы нау.-практ.конф. – Владивосток:ТИНРО-центр, 2002. – С. 39-41. 7. Саушкина Л.Н. Особенности спороношения камчатской бурой водоросли Laminaria bongardiana в антропогенной среде // Морские прибрежные экосистемы: водоросли, беспозвоночные и продукты их переработки: Тез. докл. 1–й Межд. конф. – М.: ВНИРО, 2002. – С. 35 8. Титлянов Э.А., Глебова Н.Т., Котлярова Л.С. Сезонные изменения в строении талломов Ulva fenestrata P.et R. // Экология. 1975. № 4. С. 36–41. 9. Титлянов Э.А., Колмаков П.В., Коробейников Л.С. Дневные изменения скорости видимого и потенциального фотосинтеза в течение года у некоторых бентических водорослей Японского моря // Экологические аспекты фотосинтеза морских макроводорослей. 1978. С. 136-149. 10. Усов А.И., Смирнова Г.П., Клочкова Н.Г. Полисахариды водорослей. 55. Полисахаридный состав некоторых бурых водорослей Камчатки // Биоорган. химия. 2001. Т. 27, № 6. С. 444–448. 11. Усов А.И., Смирнова Г.П., Клочкова Н.Г. Полисахариды водорослей. 58. Полисахаридный состав тихоокеанской бурой водоросли Alaria fistulosa Р. et R. (Alariaceae, Laminariales) // Известия Академии наук. Серия химическая. 2005. № 5. С. 1–5. 12. Шпакова Т.А. Рост Laminaria japonica Aresch. у юго-западного Сахалина // Изв. ТИНРО. 1985. Т. 110. C. 139–144. 13. Ярцева И.А., Возжинская В.Б., Соловьева О.Л., Каминер К.М. Физиологическая характеристика некоторых фукоидов Белого и Японского морей // Тр.ВНИРО. 1977. Т.74. С. 77-80. 14. Dische Z., Shettles L.B. A specific color reaction of methylpentoses and a spectrophotometric micromethod for their determination. // J. Biol. Chem. 1948. V. 175. P. 595-603. 15. Skriptsova A.V., Shevchenko N.M., Tarbeeva D.V., Zvyagintseva T.N. Comparative Study of Polysaccharides from Reproductive and Sterile Tissues of Five Brown Seaweeds // Marine Biotechnology. 2012. V. 14. Р. 304–311. http://link.springer.com/article/10.1007/s10126-011-9413-4 опубликовано - декабрь 2014 г.
Morphometric indices and chemical composition of the Saccharina japonica cultivated in the Japan Sea (Primorye) Natalia M. Aminina, Olga N. Guruleva Seasonal variability of the morphometric indices of the cultivated Saccharina japonica thalli is shown. Chemical composition as a whole thallus as well as its separate parts is given. Accumulation of the maximum length and weight in the two-year alga thallus occurs in June. In the same period, increasing the concentration of protein and minerals in the thalli was observed. The highest alginic acid content was detected in July and August. The most organic substances are defined in the upper part of the plate from May to June. In August, they, especially the protein, accumulate in the lower part of the plant more. The alginic acid and mannitol contents are greater in the intense biosynthesis zone along the thallus edges but the fucoidan one is in the central part of thallus. Found that the fucoidan and alginic acid amounts increase within the sporiferous tissue formation zone. Key words: brown algae; Saccharina japonica; morphometric indices; vegetation period; sporiferous tissue; chemical composition; alginic acid; mannitol; fucoidan; mineral substances; protein.
Об авторах Аминина Наталья Михайловна - Aminina Natalia Michailovna кандидат биологических наук aminina@tinro.ru Гурулева Ольга Николаевна – Guruleva Olga Nikolaevna кандидат технических наук guruleva@tinro.ru Корреспондентский адрес: Россия, 690091, г. Владивосток, пер. Шевченко, 4, ФГУП ТИНРО-центр; телефон (423) 240-13-60
ССЫЛКА НА СТАТЬЮ: Аминина Н.М., Гурулева О.Н. Морфометрические показатели и химический состав Saccharina japonica, культивируемой в Японском море (Приморский край) // Вопросы современной альгологии. 2014. № 2 (6). URL: http://algology.ru/601
Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно. При перепечатке ссылка на сайт обязательна
. На ГЛАВНУЮ
|
|||
|
| ||